часть_1_2012. Лекции по биологии в 2х кн. Ч. I. Цитология и генетика Под ред проф. Т. В. Викторовой. Уфа, 2012. 192 с., ил
 Скачать 13.87 Mb. Скачать 13.87 Mb.
|

Цитоплазматические мембраныЦитоплазматические мембраны выполняют ряд важных функций: барьерную (отграничивающую), регуляторную (осуществляют регуляцию метаболических потоков), транспортную (обеспечение избирательной проницаемости веществ путем пассивного и активного транспорта), структурную, обменную. Биологические мембраны построены в основном из липидов, белков и 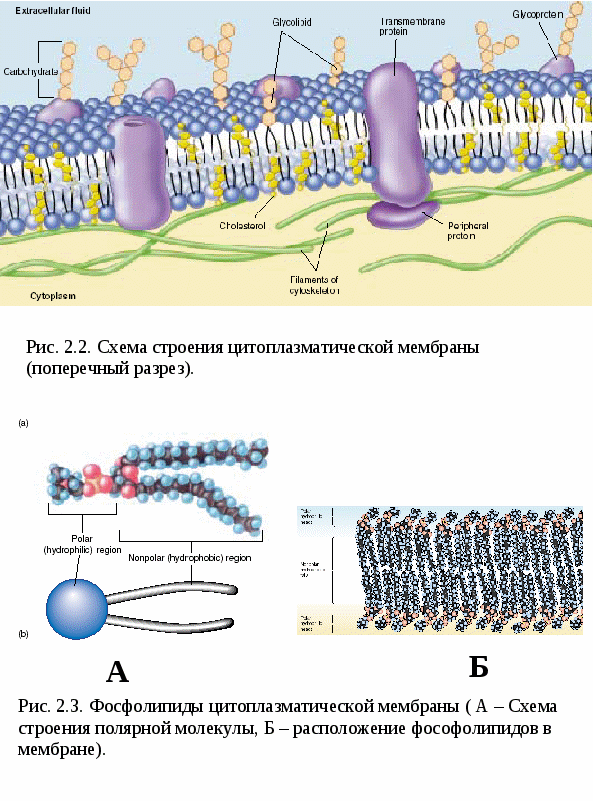 углеводов (рис.2.2). Предложено несколько моделей строения цитоплазматических мембран (модель «сэндвича» - модель Даниели и Даусона, модель Ленарда и др.). По-видимому, в зависимости от функции существует несколько типов мембран. В настоящее время принята за основу жидкостно-мозаичная модель, предложенная Сингером.-Николсоном (1972). Согласно этой модели в состав мембран входит бимолекулярный слой липидов, в который включены молекулы белков. Липиды - это водонерастворимые вещества. Они имеют полярную (заряженную) головку и длинные незаряженные (неполярные) углеводные цепи. Молекулы липидов обращены друг к другу неполярными концами, а их полярные полюса (головки) остаются снаружи, образуя гидрофильные поверхности (рис.2.3). Белки мембран можно разделить на три группы: периферические (наиболее слабо связаны с мембраной), погруженные (полуинтегральные) и пронизывающие (интегральные), формирующие поры мембраны. В функциональном отношении белки мембран подразделяются на ферментативные, транспортные, структурные и регуляторные. На внешней поверхности плазматической мембраны белковые и липидные молекулы связаны с углеводными цепями, образуя гликокаликс. Углеводные цепи выполняют роль рецепторов, клетка приобретает способность специфически реагировать на воздействия извне. Так, взаимодействие гормона со «своим» рецептором снаружи вызывает изменение структуры интегрального белка, что приводит к запусканию клеточного ответа. В частности, такой ответ может проявиться в образовании «каналов», по которым растворы некоторых веществ начинают поступать в клетку или выводятся из нее. Одна из важных функций мембраны – обеспечение контактов между клетками в составе органов и тканей. Под плазматической мембраной со стороны цитоплазмы имеется кортикальный слой и внутриклеточные фибриллярные структуры, обеспечивающие механическую устойчивость мембраны. У растительных клеток кнаружи от мембраны расположена плотная структура – клеточная оболочка, состоящая из полисахаридов (целлюлозы). Одно из важнейших свойств цитоплазмы связано со способностью пропускать в клетку или из нее различные вещества. Это необходимо для поддержания постоянства ее состава. Малые молекулы и ионы проходят через мембраны путем пассивного и активного транспорта. Пассивный транспорт происходит без затрат энергии путем диффузии, осмоса и облегченной диффузии (рис.2.4). Диффузия – 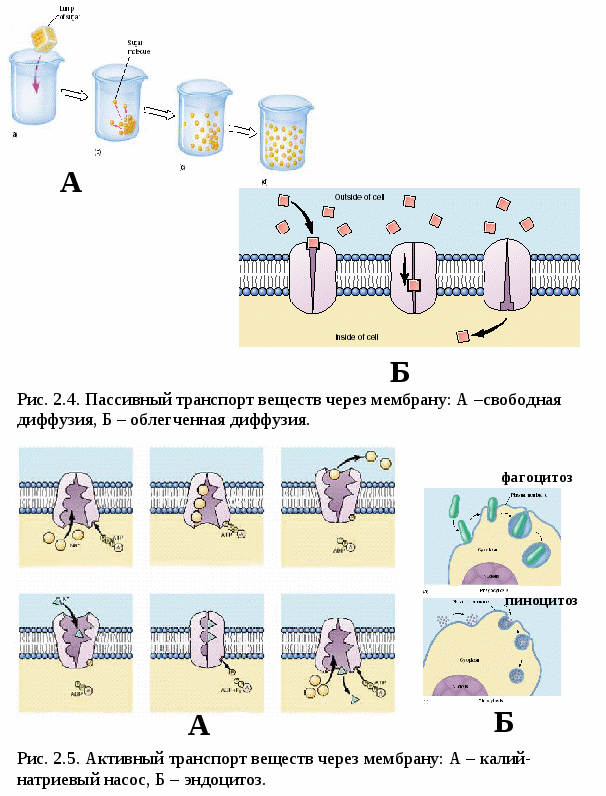 транспорт молекул и ионов через мембрану из области с высокой в область с низкой их концентрацией, т.е. по градиенту концентрации. Если вещества хорошо растворимы в жирах, то они проникают в клетку путем простой диффузии (кислород, углекислый газ). Диффузия воды через полупроницаемые мембраны называется осмосом. Вода способна проходить также через мембранные поры, образованные белками, и переносить молекулы ионов и растворенных в ней веществ. Облегченная диффузия – транспорт веществ, нерастворимых в жирах и не проходящих сквозь поры, через ионные каналы с помощью белков-переносчиков. Активный транспорт веществ через мембрану происходит с затратой энергии АТФ и при участии белков-переносчиков. Он осуществляется против градиента концентрации (так транспортируются аминокислоты, сахар, ионы калия, натрия, кальция и др.). Примером активного трантпорта может быть работа калий-натриевого насоса. Концентрация К внутри клетки в 10-20 раз выше, чем снаружи, а Na – наоборот. Для поддержания данной концентрации происходит перенос трех ионов Na из клетки на каждые два иона К в клетку. В этом процессе участвует белок в мембране, выполняющий функцию фермента, расщепляющего АТФ с высвобождением энергии, необходимой для работы насоса (рис.2.5. А). Перенос макромолекул и крупных частиц внутрь клетки осуществляется за счет эндоцитоза, а удаление из клетки - путем экзоцитоза. При эндоцитозе (рис.2.5. Б) мембрана образует впячивания или выросты, которые затем отшнуровываясь превращаются во внутриклеточные пузырьки, содержащие захваченный клеткой продукт. Этот процесс происходит с затратой энергии АТФ. Различают два вида эндоцитоза – фагоцитоз (поглощение клеткой крупных частиц) и пиноцитоз (поглощение жидких веществ). Мембрана принимает участие в выведении веществ из клетки в процессе экзоцитоза. Таким способом из клетки выводятся гормоны, белки, жировые капли и др. Цитоплазма Цитоплазма – внутреннее содержимое клетки, состоит из основного вещества, органелл и включений (рис.2.6). Гиалоплазма (цитозоль) – основное вещество цитоплазмы, заполняющее пространство между клеточными органеллами. Гиалоплазма содержит около 90% воды и различные белки, аминокислоты, нуклеотиды, ионы неорганических соединений и др. 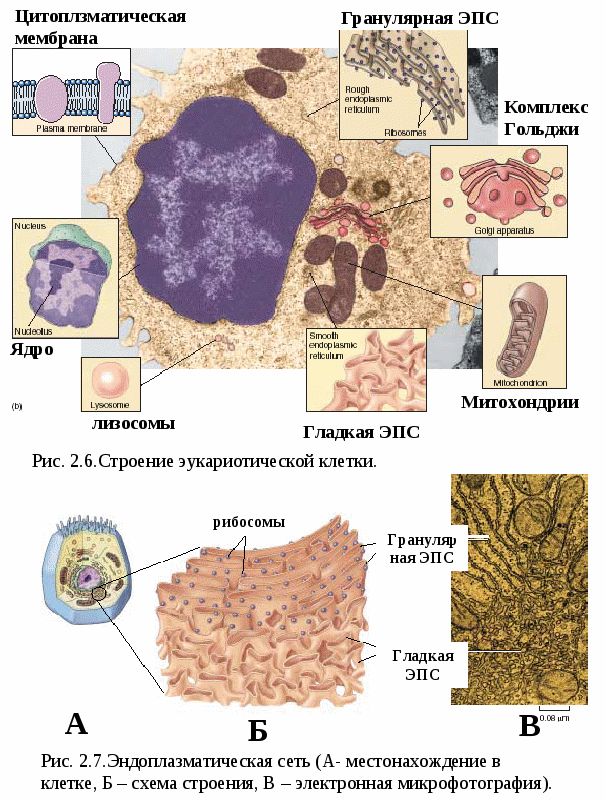 Крупные молекулы белка образуют коллоидный раствор, который может переходить из золя (невязкое состояние) в гель (вязкий). В гиалоплазме протекают ферментативные реакции, метаболические процессы, синтез аминокислот, жирных кислот. Гиалоплазма содержит множество белковых нитей – филаментов, которые пронизывают цитоплазму и образуют цитоскелет. Органеллы Органеллы (органоиды) (рис.2.6) – постоянные структуры цитоплазмы, выполняющие в клетке жизненно важные функции. В зависимости от функции различают органоиды общего и специального назначения. К органоидам специального назначения относятся микроворсинки, реснички, жгутики. Органеллы общего назначения делятся на немембранные (рибосомы, клеточный центр (центриоль), микротрубочки, микрофиламенты) и мембранные. К одномембранные органеллам относятся эндоплазматическая сеть (ретикулум), аппарат Гольджи, лизосомы, пероксисомы, вакуоли. К двумембранным органеллам относятся митохондрии и пластиды растительных клеток. Одномембранные органоиды Эндоплазматическая сеть ЭПС – это система цистерн и каналов, «стенка которых образована мембраной (рис. 2.7). Нередко цистерны имеют пузыревидные утолщения. ЭПС пронизывает цитоплазму в разных направлениях и делит ее на изолированные ячейки – компартменты. Компартментализация способствует пространственному разделению веществ и процессов в клетке. ЭПС выполняет синтетическую и транспортную функции. Если на поверхности мембран каналов ЭПС располагаются рибосомы, она называется гранулярной или шероховатой, если рибосом нет – гладкой. Функции ЭПС: 1) биосинтез белков (гранулярная ЭПС), жиров и углеводов (гладкая ЭПС), 2) транспортировка всех веществ в клетке, 3) компартментализация цитоплазмы (разделение на отсеки), 4) участие в образовании мембран цитоплазмы. Отчленяющиеся от ЭПС пузырьки представляют исходный материал для других одномембранных органелл: аппарата Гольджи, лизосом, вакуолей. Аппарат Гольджи (пластинчатый комплекс) Аппарат Гольджи (в честь К. Гольджи, который обнаружил органеллу в 1898 г.) обычно расположен около клеточного ядра. 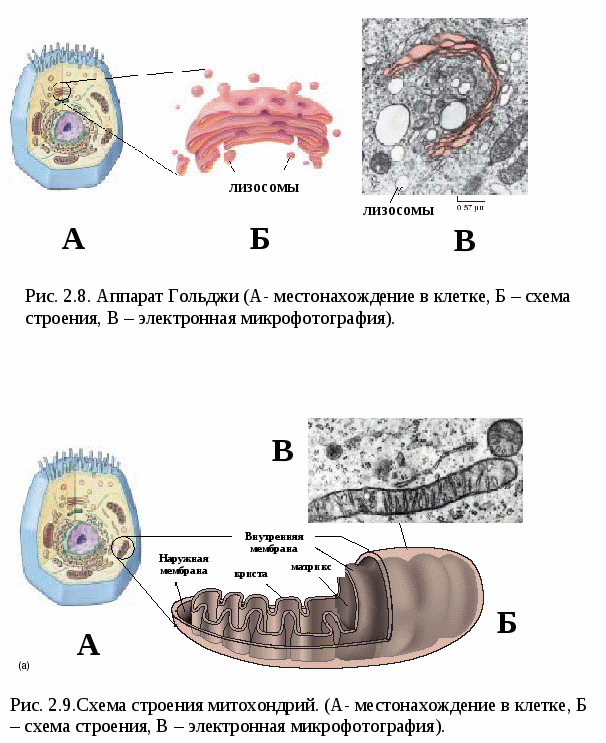 Основным элементом органеллы является мембрана, образующая уплощенные цистерны – диски, которые располагаются друг над другом (4-6). Края цистерн переходят в трубочки, от которых отчленяются пузырьки, транспортирующие заключенное в них вещество к месту его потребления (лизосомы, вакуоли) (рис. 2.8). Поэтому наиболее крупные аппараты Гольджи находятся в секретирующих клетках. Диски-цистерны формируются из пузырьков, отпочковывающихся от гладкой ЭПС. Функции: секреция веществ, их сортировка и упаковка, образование комплексных соединений, формирование лизосом. Лизосомы Лизосомы (от греч. Лизис – разрушение, сома – тело) – пузырьки больших или меньших размеров, заполненные ферментами (протеазами, липазами, нуклеазами). Лизосомы образуются в ЭПС и аппарате Гольджи. Основная функция лизосом – внутриклеточное расщепление и переваривание веществ, поступивших в клетку и удаление их из клетки. Выделяют первичные и вторичные лизосомы. Пузырьки с набором ферментов, отделившиеся от цистерн аппарата Гольджи, называют первичными лизосомами. Они участвуют во внутриклеточном пищеварении. Если первичные лизосомы сливаются с фагоцитарными и пиноцитарными вакуолями, образуются вторичные лизосомы. В них происходит переваривание веществ, поступивших в клетку путем эндоцитоза. Продукты переваривания поглощаются клеткой, а лизосомы, содержащие нерасщепленные материал, называются остаточными тельцами, которые путем экзоцитоза выводятся наружу. Вакуоли. Содержатся в цитоплазме клеток растений. Образуются из мелких пузырьков, отщепляющихся от ЭПС. В вакуолях запасается вода, питательные вещества (белки, сахара), откладываются пигменты. Вакуоли поддерживают тургор (давление) в клетке. Некоторые вакуоли напоминают лизосомы. Пероксисомы – органеллы, освобождающие клетки от перекисей, накапливающихся вследствие неферментативного окисления жирных кислот. Двумембранные органоиды Митохондрии Митохондрии – это структуры округлой или палочковидной формы (рис. 2.9). Обычно митохондрии скапливаются в тех участках, где велика потребность в АТФ (скелетные мышцы, сердце). Состоит из двух мембран. Наружная мембрана гладкая, внутренняя образует многочисленные складки – кристы. На кристах митохондрий расположены сферические тельца на ножках – АТФ-сомы. Митохондрии называют энергетическими 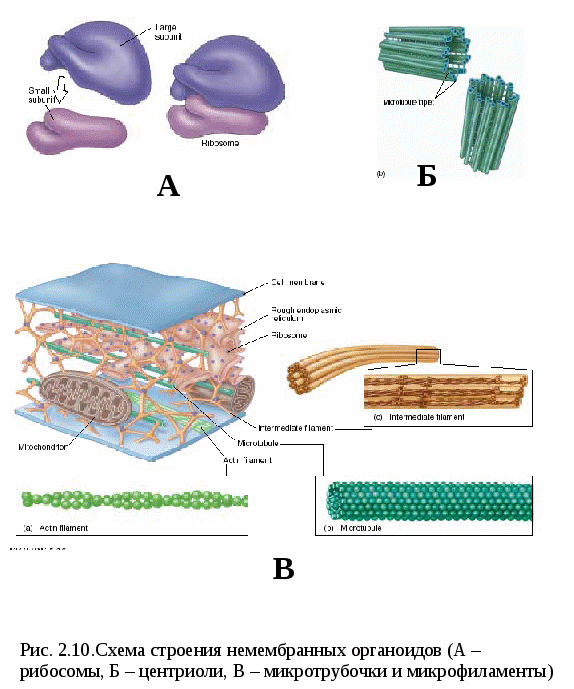 станциями клетки. В них происходит окисление органических веществ, благодаря чему освобождается заключенная в них энергия. Митохондрии содержат три группы ферментов: цикла Кребса (катализируют окислительно-восстановительные реакции, внутренний матрикс), тканевого дыхания (кристы) и окислительного фосфорилирования (АТФ-сомы). Энергия АТФ используется: 1) для биосинтеза веществ (50%), 2) для транспортировки (30-40%), 3) для механической работы – сокращение мышц, 4) для деления клеток, 5) рассеивается в виде тепла. Пластиды Это двумембранные органеллы, присутствующие в растительных клетках. Различают три вида пластид: хлоропласты, хромопласты, лейкопласты. Немембранные органоиды Рибосомы На рибосомах осуществляется соединение аминокислот в полипептидные цепочки (синтез белка). Каждая рибосома состоит из двух частей: малой и большой субъединиц. Объединение их происходит в присутствии мРНК (рис.2.10 А). Клеточный центр Органоид характерен для животных клеток. Располагается около ядра. Состоит из парных центриолей, расположенных перпендикулярно, и центросферы (рис.2.10 Б). Центриоль имеет вид полого цилиндра, стенка которого образована 27 микротрубочками (9 триплетов). В функцию центриолей входит образование нитей митотического веретена деления, которые также образованы микротрубочками. Центриоли поляризуют процесс деления клеток, обеспечивая расхождение сестринских хроматид. Микротрубочки и микрофиламенты Микротрубочки – тончайшие трубочки, стенки которых образованы белком тубулином (рис.2.10 В). Микрофиламенты – тонкие белковые нити, состоят из белка актина. Участвуют в образовании нитей веретена деления и цитоскелета. Органоиды специального назначения. Реснички и жгутики – органеллы передвижения. Представляют собой тонкие цилиндрические выросты цитоплазмы, покрытые плазматической мембраной. Жгутики отличаются от ресничек длиной. У основания ресничек и жгутиков лежат базальные тельца. 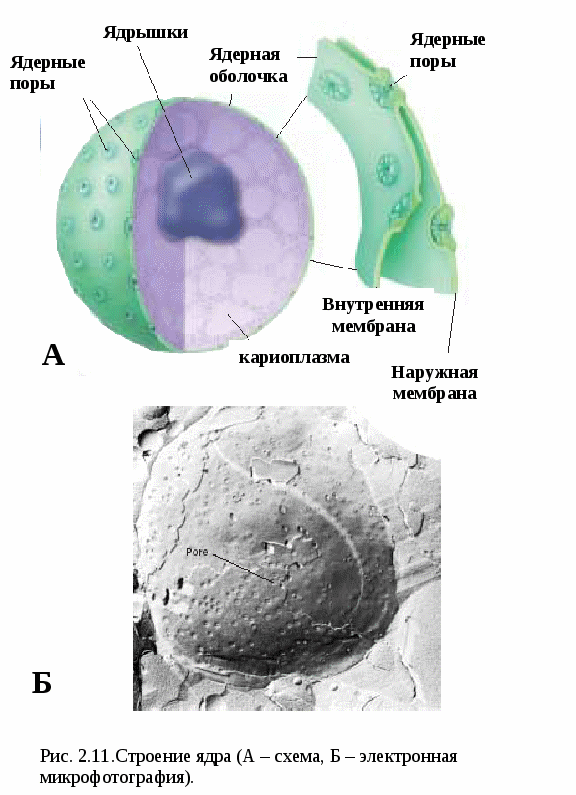 Структура клеточного ядра Ядро является постоянным структурным компонентом всех клеток высших растений и животных. Оно присутствует во всех эукариотических клетках за исключением зрелых эритроцитов крови человека и некоторых животных. Биологическое значение ядра заключается в регуляции всех жизненно-важных функций клетки и в передаче наследственной информации. В ядре хранится наследственная информация, заключенная в ДНК, которая при делении клетки передается дочерним клеткам. Ядро определяет специфичность белков, синтезируемых данной клеткой. В ядре синтезируется РНК. Ядро имеет ядерную оболочку, отделяющую его от цитоплазмы, кариоплазму (ядерный сок), хроматин, одно или несколько ядрышек (рис. 2.11). Ядерная оболочка образована двумя мембранами: внешней и внутренней. Каждая мембрана соответствует элементарной мембране и имеет трехслойное строение (бимолекулярный слой липидов, поверхостный аппарат – гликокаликс, внутренний опорно-сократительный аппарат). Промежуток между мембранами называется перинуклеарным пространством. Наружная ядерная мембрана имеет контакт с внутриклеточными мембранами, в частности, может переходить в мембраны ЭПС. Некоторые ученые считают эту мембрану производной ЭПС. На наружной мембране с внешней стороны находятся рибосомы, синтезирующие специфические белки. Внутренняя мембрана связана с хромосомным материалом ядра. Ядерная оболочка пронизана большим количеством пор (поровый комплекс) диаметром 30-40 нм до 120 нм. Поры играют важную роль в переносе веществ в цитоплазму и из нее. Число пор подвержено значительным вариациям, оно зависит от размеров ядер и функциональной активности клетки. Поры занимают до 10-15% поверхности всего ядра. Поровый комплекс представляет собой участок, в котором наружная и внутренняя ядерные мембраны соединяются между собой. Однако поры не являются пустыми пространствами. Они имеют сложную гетерогенную белковую структуру (белковые глобулы). В порах расположен канал из белковых глобул, через который в цитоплазму транспортируется мРНК. Ядерный сок (кариоплазма) – внутреннее содержимое ядра, представляет собой раствор белков, нуклеотидов, ионов, более вязкий, чем гиалоплазма. В кариоплазме находятся ядрышки и хроматин. Ядерный сок обеспечивает нормальное функционирование генетического материала. Хроматин представляет собой дезоксирибонуклеопротеин. Это комплекс молекулы ДНК с гистоновыми белками. Хроматин в электронный микроскоп выявляется в виде тонких нитей, глыбок и гранул. В процессе митоза хроматин удваивается и конденсируется (спирализуется), образуя, таким образом, хорошо видимые при окраске структуры – хромосомы. ДНК, входящая в состав хроматина, представляет собой двухцепочечную спиральную молекулу, которая укомплектована в хромосомы в комплексе с белками. Такая структура называется дезоксирибонуклеопротеидом – ДНП. На долю белков приходится 65% массы хромосом. Все хромосомные белки разделяются на 2 группы: гистоны (основные) 40% и негистоновые (кислые) белки 20%. Ядрышки – непостоянные образования, они исчезают при делении клеток и восстанавливаются после окончания деления. Ядрышки образованы определенными участками хромосом (т.н. ядрышковые организаторы), содержащими рибосомные гены. Т.о. в ядрышках происходит формирование субъединиц рибосом, которые затем через поры выходят из ядра в цитоплазму. Глава 3 Хроматин: структура, функции, уровни укладки. Жизненный и митотический цикл клетки. Митоз. Другие способы репродукции соматических клеток (амитоз, эндомитоз, эндоредупликация) Хроматин представляет собой дезоксирибонуклеопротеин. Это комплекс молекулы ДНК с гистоновыми белками. Хроматин в электронный микроскоп выявляется в виде тонких нитей, глыбок и гранул. В процессе митоза хроматин спирализуется и образует хорошо видимые окрашенные структуры – хромосомы. ДНК, входящая в состав хроматина, представляет собой двухцепочечную спиральную молекулу, которая укомплектована в комплексе с белками. Такая структура называется дезоксирибонуклеопротеидом – ДНП. На долю белков приходится 65% массы хромосом. Все хромосомные белки разделяются на 2 группы: гистоны (основные) 40% и негистоновые (кислые) белки 20%. Гистоны играют особую роль в структурной организации ДНП. Гистоны имеют «+» заряд, что обусловлено высоким содержанием в них 3х основных аминокислот: аргинина, лизина и гистидина. Они обладают высоким сродством к молекуле ДНК, которая имеет «» заряд и образует с ней прочные структурные комплексы. Это препятствует считыванию заключенной в молекуле ДНК биологической информации (регуляторная роль гистонов). Число фракций негистоновых белков превышает 100. Среди них ферменты синтеза и процессинга РНК, редупликации и репарации ДНК. Длина интерфазных хромосом (хроматина) в 1 клетке человека равна примерно 2 м (2.000.000 мкм). При переходе в метафазное состояние нить ДНК уменьшается в линейном размере почти в 8000 раз!, а диаметр увеличивается в 500-600 раз что свидетельствует о громадных масштабах физического преобразования. Рассмотрим основные закономерности поперечной и продольной укладки хромосом. Выделяют 4 уровня укладки ДНК в хроматине: 1) нуклеосомный; 2) нуклеомерный (соленоид); 3) хромомерный (петлевой); 4) хромонемный (рис.3.1). Первый уровень укладки молекулы ДНК - нуклеосомная нить. Наиболее типичными структурами хроматина, выявляемыми в электронном микроскопе, являются нити диаметром 1030 нм. Эти нити состоят из ДНК и гистонов (Н2А; Н2В; Н3 и Н4), формируя нуклеогистон. Гистоны образуют белковые тела коры (corсердцевина), 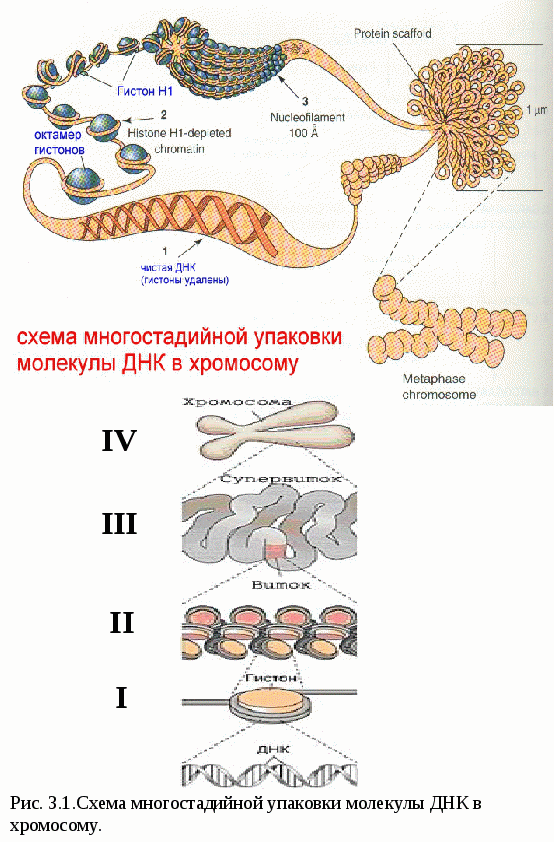 состоящие из 8 молекул (по 2 каждого гистона). Молекула ДНК образует комплекс с белковыми корами, спирально накручиваясь на них и образуя нуклеогистон. На один нуклеогистон приходится 200 пн. Второй уровень укладки нуклеомерный «соленоид». Обеспечивается гистоном Н1, который сближает белковые коры. В результате образуется более компактная фигура, возможно, построенная по типу «соленоида» формируется элементарная хромосомная фибрилла. I и II уровни укладки характерны для интерфазных хромосом – глыбок хроматина. Третий уровень укладки петлевой хромомерный. Обусловлен укладкой элементарной хроматиновой фибриллы в петли. Соответствует ранней прометафазной хромосоме. В образовании петлевых структур, повидимому, принимают участие негистоновые белки, которые способны узнавать специфические участки молекулы ДНК, отдаленные друг от друга на расстояние в несколько тысяч нуклеотидов, и сближать их с образованием петель из расположенных между ними фрагментов хроматиновой фибриллы. 1 петля соответствует 2080 тысячам пар нуклеотидов. Возможно, каждая петля является функциональной единицей генома. Четвертый уровень укладки хромонемный. (соответствует метафазной хромосоме). Наиболее простым и приемлемым является признание спиральной укладки каждой хроматиды. У самых крупных хромосом человека (1 и 2) 14 15 таких витков. У мелких 24 витка. Набор хромосом в соматических клетках данного вида организмов называется кариотипом. Кариотип – это видоспецифический признак, характеризующийся определенным числом, строением и генетическим составом хромосом. Этот термин был введен в 1924 г. советским генетиком Левитским. Понятие кариотип подразумевает диплоидный набор хромосом. У каждой хромосомы имеется гомолог, т.е. хромосома, с таким же набором генов. Один набор хромосом – от матери, другой – от отца. Если число хромосом в гаплоидном наборе (в половых клетках) обозначить как n, то формула кариотипа будет 2n. Изучение полного набора хромосом называется кариотипированием. Важность постоянства кариотипа определена в 4-х правилах хромосом: постоянства, парности, индивидуальности и непрерывности. Нормальный кариотип человека включает 46 хромосом, или 23 пары: из них 22 пары – аутосомы (одинаковые по строению и набору генов) у 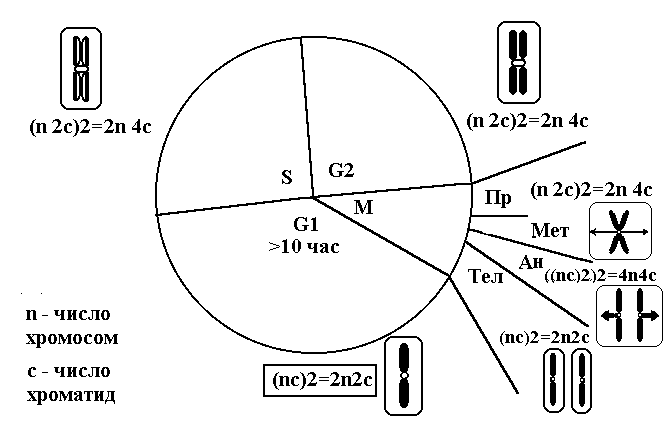 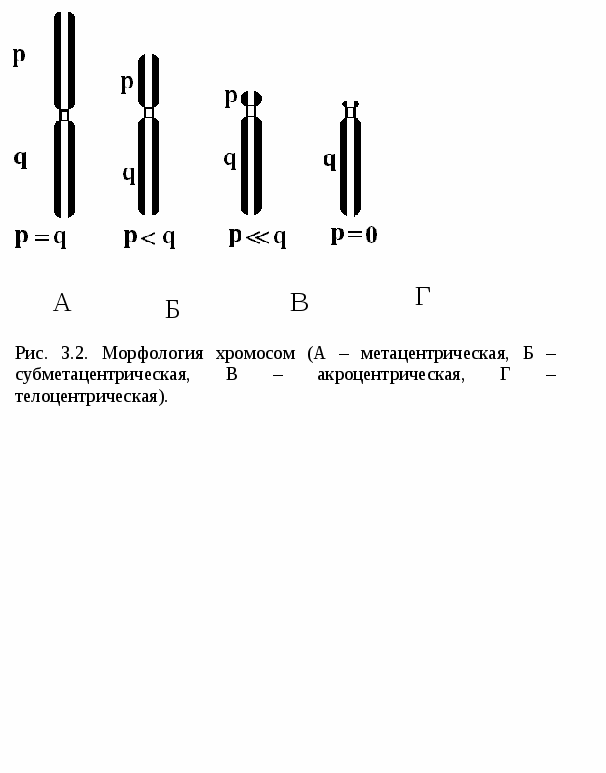 Рис. 3.3. Клеточный цикл представителей разного пола и 1 пара половых хромосом (гетерохромосомы): ХХ у женщин и XY – у мужчин. Для изучения кариотипа клетки изучают на стадии метафазы, когда хромосомы максимально конденсированы. Именно на этой стадии можно наиболее точно определить морфологию каждой хромосомы. Морфология хромосом В структуре хромосом выделяют: длинное плечо, короткое плечо, центромера, II перетяжка – ЯОР. По месту расположения II перетяжки различают метацентрические, субметацентрические, акроцентрические, телоцентрические хромосомы (рис. 3.2). Клеточный цикл и митотический цикл клетки (МЦК) Особое значение для нормальной жизнедеятельности клеток организма имеет поддержание постоянства кариотипа в клетках организма, которое обеспечивают два важных механизма: 1. воспроизведение хромосом, 2. распределение хромосом при делении клеток. В основе воспроизведения хромосом лежат процессы их самоудвоения, т.е. репликация ДНК. Правильное и точное распределение хромосом при делении клеток обеспечивается митозом. Эти два механизма в совокупности образуют митотический цикл клетки (МЦК). В отличие от МЦК клеточный цикл включает период G0, в течение которого происходит рост, дифференцировка и функционирование клеток, иногда специализация с потерей способности к делению и естественная гибель. МЦК – это совокупность процессов, происходящих в клетке от одного деления до следующего и заканчивающихся образованием из одной материнской двух новых дочерних клеток с неизменным набором хромосом. В МЦК условно можно выделить два периода: собственно митоз (деление) и интерфазу. В интерфазе различают 3 периода: пресинтетический (G1-период), синтетический (S-период), и постсинтетический (G2-период). В пресинтетическом периоде клетки имеют диплоидный набор хромосом (2n), причем, каждая хромосома состоит из 1 хроматиды (рисунок). Формула хромосом 2n2c. В синтетическом периоде происходит репликация ДНК. После окончания S-периода каждая хромосома состоит уже не их 1, а из 2 сестринских хроматид (образуются дочерние хромосомы). Формула хромосомного набора, соответственно, приобретает вид 2n4c (на 2 хромосомы теперь приходится 4 хроматиды) (рисунок). В постсинтетическом клетка готовится к делению. Формула хромосом остается прежней – 2n4c. 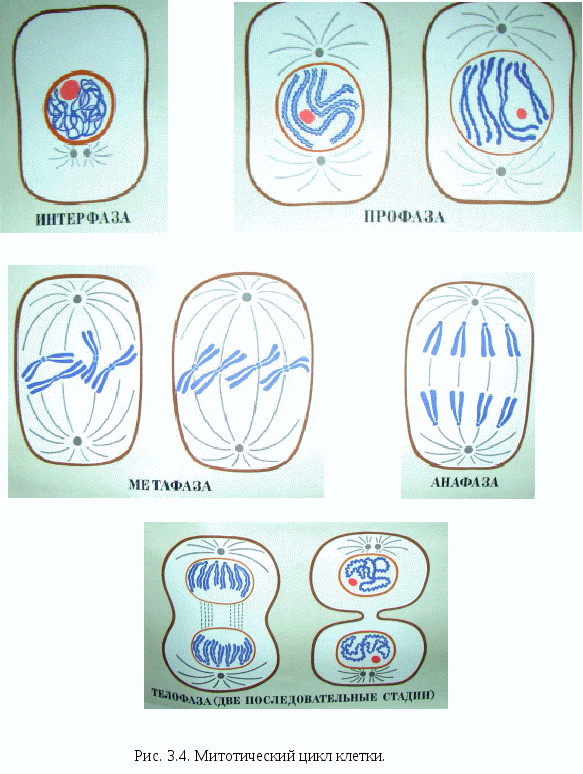 За интерфазой следует непосредственно деление клеток - митоз, в котором различают два этапа: Деление ядра клетки – кариокинез; Деление цитоплазмы клетки – цитокинез. Митоз Митоз состоит из 4 фаз: 1 – профаза (иногда выделяют прометафазу); 2 – метафаза; 3 – анафаза; 4 – телофаза. Каждая предыдущая фаза обуславливает переход к следующей (рис. 3.4). Биологическое значение митоза Заключается в точном идентичном распределении сестринских хроматид (или дочерних хромосом) между дочерними клетками. В результате этого деления из одной материнской клетки образуются две генетически равноценные дочерние клетки, которые имеют хромосомный набор, по количеству и качеству точно идентичный материнской клетке. Так поддерживается постоянство кариотипа (т.е. набора хромосом) в поколениях клеток. Разновидностями митоза являются амитоз, эндомитоз и эндоредупликация. Амитоз – прямое деление цитоплазмы без формирования веретена деления (у человека в норме встречается в клетках печени). При эндомитозе удвоение ДНК и деление центромер и хромосом в анафазе не сопровождается делением ядра и клетки, в результате чего набор хромосом в ядре удваивается. Это приводит к полиплоидии, когда число хромосом увеличивается пропорционально 2n: 2n (норма, диплоид) – 4n (тетраплоид) и т.д. В результате эндоредупликации происходит удвоение ДНК без деления центромер, в результате чего число хроматид в одной хромосоме увеличивается пропорционально 2с: 2с -4с-8с-16с и т.д. Образуются политенные (от лат. tensis – нить) хромосомы, содержащие до 1000 нитей хроматина, объединенных одной центромерой. Нервно-гуморальные факторы регуляции митоза Среди факторов нервно-гуморальной регуляции митоза особое место отведено двум ключевым белкам: циклины и циклин-зависимые киназы. Циклины активируют CDK, что является сигналом к удвоению хромосом и делению клетки. В процессе митотического деления каждая клетка получает полный двойной комплект хромосом. Если бы образование половых клеток шло таким же путем, то после оплодотворения (слияния женской и мужской гаметы) число хромосом всякий раз удваивалось бы. На самом деле этого не происходит. Каждому виду свойственно определенное число и определенный набор хромосом (кариотип). Это возможно лишь при условии, если при образовании клеток будет происходить редукция, т.е. уменьшение, числа хромосом вдвое. 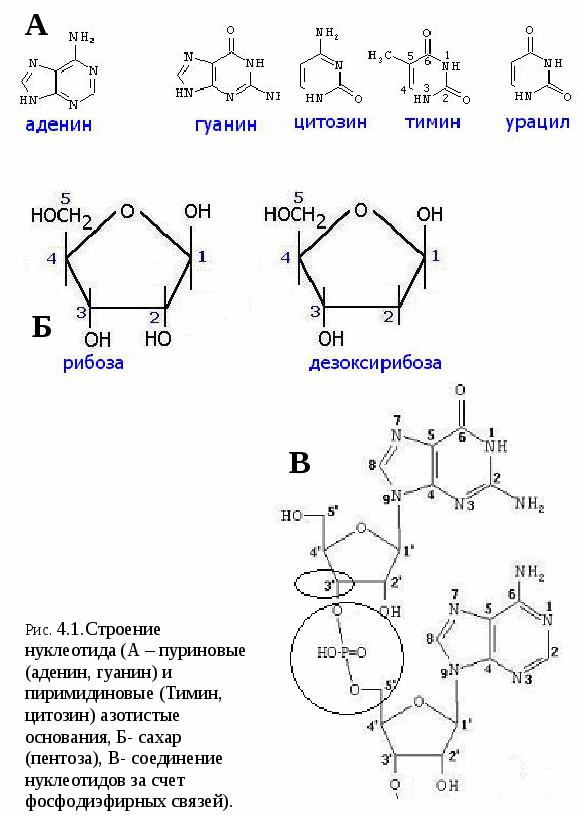 Глава 4 Генный уровень организации наследственного материала. Химическая и структурная организация нуклеиновых кислот (ДНК и РНК). Репликация и репарация ДНК. Структура генов прокариот и эукариот. Генетический код и его свойства Исследования, направленные на выяснение химической природы наследственного материала показали, что материальным субстратом генов являются нуклеиновые кислоты – ДНК и РНК. Нуклеиновые кислоты – это макромолекулы, т.е. отличаются большой молекулярной массой. В структурной организации молекулы ДНК можно выделить 3 уровня: Первичная структура – полинуклеотидная цепь Вторичная структура – две комплементарные друг другу и антипараллельные полинуклеотидные цепи Третичная структура – трехмерная спираль Первичная структура ДНК ДНК – это полимер, состоящий из мономеров – нуклеотидов. Каждый нуклеотид включает три основные компонента: азотистое основание (аденин, гуанин, цитозин, тимин, урацил), сахар (дезоксирибоза или рибоза), остаток фосфорной кислоты (рис. 4.1). Соединение нуклеотидов в макромолекулу нуклеиновой кислоты происходит путём взаимодействия фосфата одного нуклеотида с гидроксилом д.р. так, что между ними устанавливается фосфодиэфирная связь. В результате образуется полинуклеотидная цепь. Важно отметить, что сборка полипептидной цепи осуществляется строго в одном направлении, а именно, путем присоединения фосфатной группы, расположенной в 5’-положении последующего нуклеотида к 3’гидроксильной группе предыдущего нуклеотида. Вторичная структура ДНК В 1953 г Уотсон и Крик представили модель 3-х мерной молекулы ДНК (рис. 4.2). За это открытие в последующем они были удостоены Нобелевской премии. Ученые показали, что особенностью II структурной организации ДНК является то, что в ее состав входят 2 полипептидные цепи, связанные между собой особым образом – путём образования водородных связей между азотистыми основаниями по принципу комплементарности: пурин связывается только с пиримидином, т.е. Аденин (А) может связываться только с Тимином (Т), а Гуанин (Г, англ. G) – только с Цитозином (Ц, англ. C) и наоборот. При этом между А и Т образуются 2 водородные 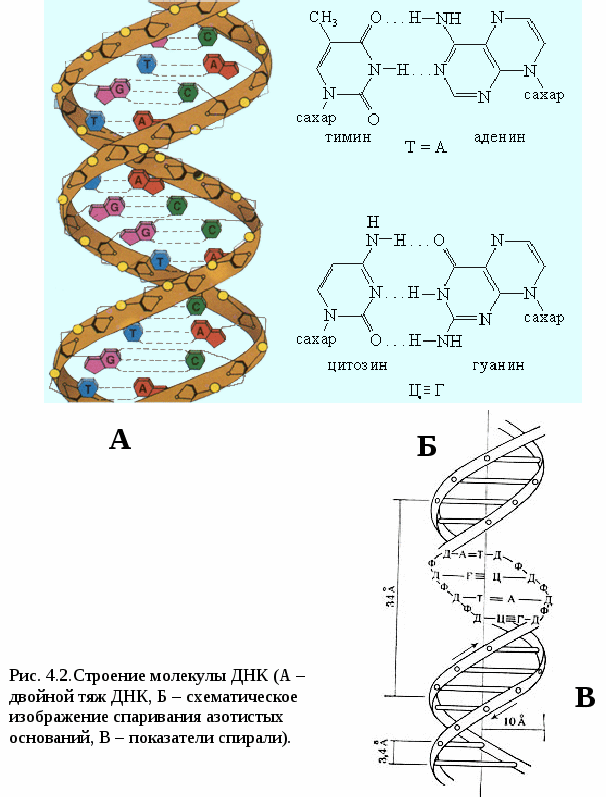 связи, а между Г и Ц – 3. Благодаря комплементарности соединения в цепь число пуринов в молекуле ДНК всегда равно числу пиримидинов, т.е. A + Г = T + Ц – это правило Чаргаффа (1951 г.) Хотя водородные связи между парами оснований относительно слабы, каждая молекула ДНК содержит приблизительно 3,3 млрд. пар, так что в физиологических условиях (Т0 , рН) цепи никогда не разрываются. Сахаро-фосфатный остов находится по периферии молекулы ДНК, а пуриновые и пиримидиновые основания – внутри. Другой важной особенностью молекулы ДНК является антипараллельность двух составляющих её цепей, т.е. 5’– конец одной цепи соединяется с 3’-концом другой. Третичная структура ДНК Данные рентгеноструктурного анализа показали, что молекула ДНК образует правозакрученную спираль диаметром 2 нм; длиной шага – 3,4 нм. В каждый виток входит 10 п.н. ДНК эукариот неоднородна по функциональной значимости и может быть подразделена на 3 класса: 1. Повторяющиеся последовательности (не транскрибируются) встречаются в геноме до 1млн. раз. 2. Умеренно повторяющиеся последовательности (встречаются в геноме 10² - 10³ раз). Это гены тРНК и белков, входящих в состав рибосом, хроматина и рРНК. 3. Уникальные участки с неповторяющимися сочетаниями нуклеотидов. У человека уникальные участки ДНК составляют не более 10-15% от общей длины молекулы ДНК. Уникальные участки ДНК являются структурной основой большинства генов человека, в которых закодирована информация о первичной структуре полипептида. Отсюда следует, что жизнедеятельность организма обусловлена, в основном, функциональной активностью уникальных участков, т.е. генов. Главная функция ДНК заключается в том, что она предназначена для хранения и передачи наследственной информации в клетках про- и эукариот. У вирусов эту функцию выполняет РНК. Основные отличия ДНК от РНК
2. По строению она сходна с одной из цепей ДНК, только вместо Тимина, входящего в состав молекулы ДНК в молекуле РНК присутствует Урацил (У) (пиримидиновый нуклеотид). 3. Между ДНК и РНК существуют различия в характере углевода: в ДНК- дезоксирибоза, в РНК – рибоза. В отличие от ДНК, содержание которого в клетке постоянно, содержание в них РНК сильно колеблется и зависит от интенсивности синтеза белка. Виды РНК 1.Транспортная (тРНК). Состоит из 75-95 нуклеотидов. Осуществляет перенос аминокислот в рибосомы, т.е. к месту синтеза полипептида. 2.Рибосомная (рРНК). Включает 3-5 тысяч нуклеотидов. р РНК составляет существенную часть структуры рибосомы. 3.Информационная, или матричная (иРНК, или мРНК) – осуществляет перенос информации о структуре белка от ДНК к месту синтеза белка в рибосомах. Важно отметить, что все виды РНК синтезируются по матрице ДНК! Нуклеиновые кислоты отличают от всех других макромолекул два удивительных свойства.
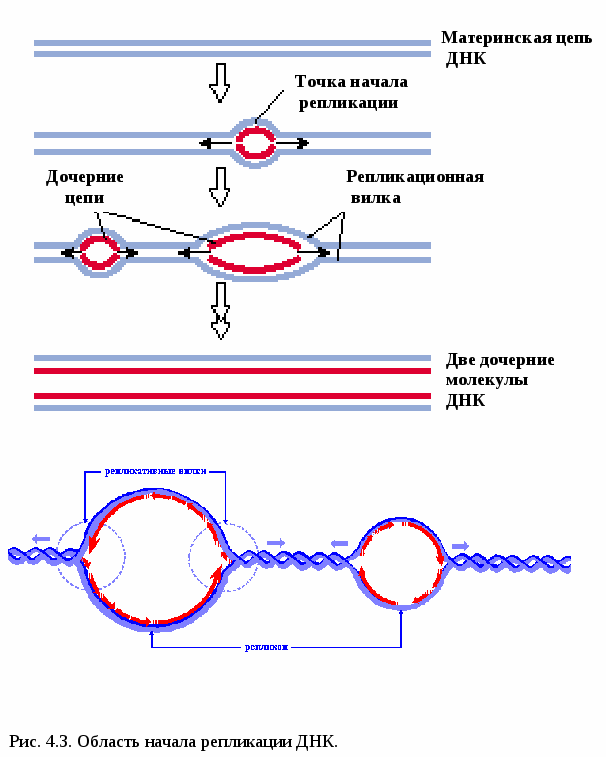
Рассмотрим эти два свойства молекулы ДНК. Самовоспроизводство - репликация молекулы ДНК Артур Корнберг в 1956 провел уникальные опыты. Он синтезировал ДНК in vitro, т.е. в пробирке. Репликация (удвоение) ДНК происходит в S-периоде клеточного цикла (S - sinthesis). В результате из одной молекулы ДНК образуются две идентичные двойные спирали. Каждая из двух цепей материнской молекулы служит матрицей для «дочерей». После репликации в обеих двойных спиралях одна цепочка – материнская, другая- дочерняя. Такой способ удвоения молекул ДНК называется полуконсервативным. Репликация ДНК начинается в строго определенных точках, имеющих уникальные последовательности длиной около 300 п.н., которую узнают специальные инициирующие белки. Эти участки называются точками начала репликации - (Ori – Origine - источник). У прокариот имеется одна точка Ori, у эукариот их много (рис. 4.3). 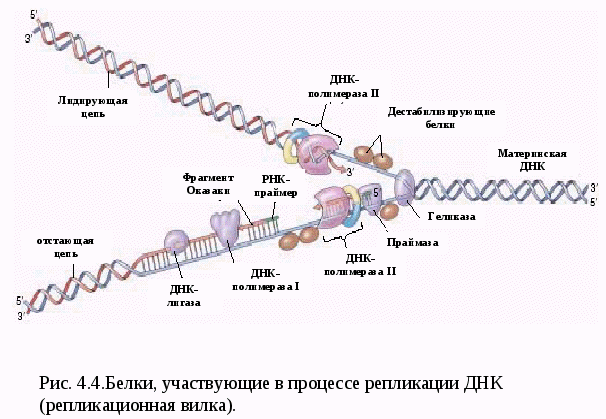 Двойная спираль ДНК раскручивается и образуется репликационная вилка. Участок молекулы ДНК от точки начала одной репликации до точки начала другой репликации называется репликоном. В процессе репликации ДНК принимает участие множество ферментов (рис.4.4). 1. ДНК-геликаза разрывает водородные связи и двойная спираль расплетается. 2. Дестабилизирующие белки соединяются с одноцепочечной ДНК и фиксируют ее. 3. В результате раскручивания цепи ДНК возникает суперспирализация. ДНК-топоизомераза – фермент, который разрывает одну из цепей ДНК и дает ей возможность свободно вращаться вокруг другой цепи. Это снимает напряжение в спирали ДНК. 4. Начало репликации активируется короткими фрагментами РНК (100-200 нуклеотидов), которые называются РНК – затравкой или РНК-праймерами. 5.ДНК- полимераза синтезирует цепь ДНК от места присоединения РНК- затравки. Новая цепь синтезируется в направлении от 5’ к 3’ , т.к. 5 конец последующего нуклеотида с остатком фосфорной кислоты присоединяется к 3’ОН-концу (гидроксильная группа) уже синтезированного участка ДНК. Синтез ДНК происходит полунепрерывно, поскольку различают лидирующую (ведущую цепь) и отстающую цепь. На отстающей цепи синтезируются фрагменты Оказаки длиной 1000-2000 нуклетидов. 6. ДНК-лигаза соединяет (сшивает) отдельные фрагменты Оказаки. За один митотический цикл ДНК клетки полностью реплицируется только один раз. Пока полностью не закончится репликация ДНК, не происходит деления клетки. Репликация ДНК и про- и эукариот в основных чертах протекает сходно. Однако скорость репликации у эукариот составляет около 100 нуклеотидов в секунду, что на порядок ниже, чем у прокариот (1000 н/сек). Это необходимо для высокой точности репликации ДНК (1 ошибка на 100.000 нуклеотидов). Но благодаря наличию множества точек Ori репликация ДНК у эукариот происходит достаточно быстро. Репарация ДНК Точность репликации ДНК у эукариот поддерживается несколькими механизмами. Во-первых, это самокоррекция, т.е. отщепление ошибочно присоединенного нуклеотида. Этот процесс контролирует ДНК-полимераза и частота ошибок уменьшается в 10 раз (1 ошибка на 10.000 нуклеотидов). Во-вторых, в организме действует система репарации, т.е. восстановления повреждений ДНК и исходной последовательности 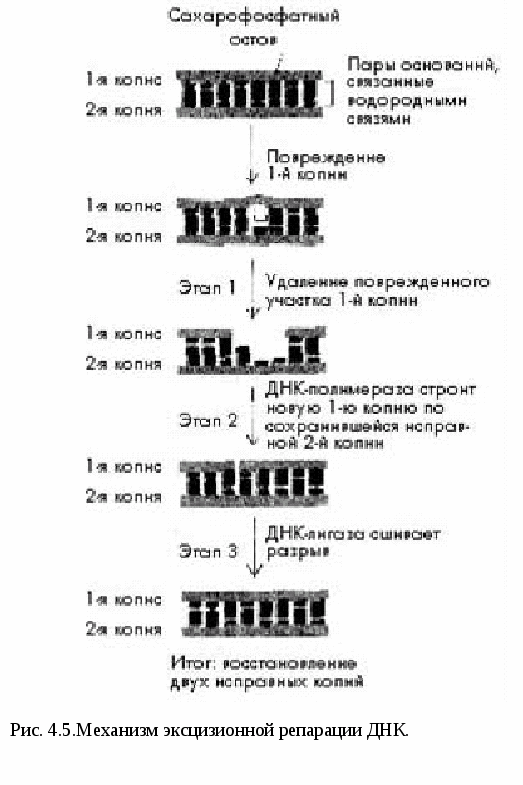 нуклеотидов. Наиболее распространенным механизмом репарации ДНК является эксцизионная репарация – вырезание ошибочных нуклеотидов. Механизм эксцизионной репарации ДНК(рис.4.5) .
Итак, молекула ДНК- носитель всей генетической информации в клетке. Структура ДНК - набор нуклеотидов. Следовательно, структурной единицей генов являются нуклеотиды (ATGC). Структурной единицей белков являются аминокислоты. Первым белком, для которого была определена последовательность аминокислот, был белок инсулина. В молекуле ДНК зашифрована информация о последовательности аминокислот в полипептидах (или белках) с помощью биологического (или генетического) кода. Т.о. Генетический код – это запись в уникальных участках молекулы ДНК информации о структуре белков и полипептидов. Каким же образом в молекуле ДНК зашифрована информация о структуре белка? Ответ на этот вопрос был дан в 1961 году Фрэнсисом Криком. Ф. Крик и его коллеги предположили, что информация должна быть выражена через блоки – кодоны. Они предположили, что кодоны должны включать не менее 3-х нуклеотидов. Почему? В природе обнаружено 20 различных аминокислот, из которых комплектуются все белки. Для того, чтобы зашифровать 20 вариантов аминокислот, генетический код должен включить как минимум 3 нуклеотида, т.к. из двух нуклеотидов можно скомбинировать только 42 =16 вариантов, а из трех нуклеотидов – 43 = 64 варианта. Полная расшифровка генетического кода проведена в 60-х годах XX века. Оказалось, что из 64 возможных вариантов триплетов 61 кодирует различные аминокислоты, а 3 являются бессмысленными, или STOP-кодонами: UAG, UAA, UGA кодонами, на которых прекращается считывание наследственной информации (рис. 4.6). Свойства генетического кода
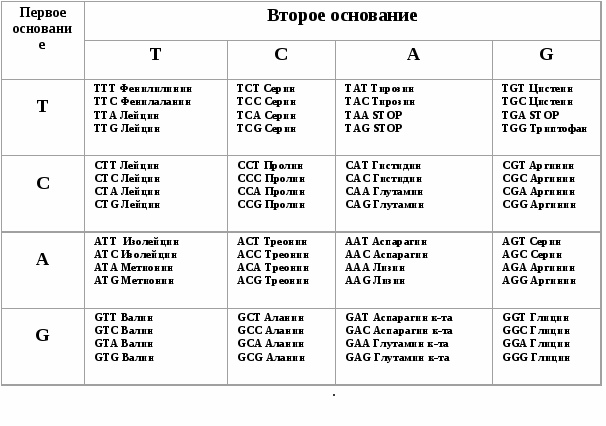 Рис. 4.6. Генетический код. Триплетные кодоны ДНК и соответствующие им аминокислоты триплетами. Это имеет очень важное значение, поскольку замена нуклеотида не всегда может приводить к замене аминокислоты). Например, аминокислоту валин кодируют три триплета: GTT, GTC, GTA, GTG.
Рассмотрим условные примеры: Норма: 123 123 123 123 … Делеция (утрата): 123 231 231 231 … (произошел сдвиг рамки считывания). Вставка: 123 112 312 312 312 …(произошел сдвиг рамки считывания). Классификация генов Все гены по функциям подразделяются на структурные и функциональные (рис.4.7).
Все клетки многоклеточного организма, возникая из зиготы путем митоза, получают полноценный набор генетической информации. Несмотря на это, они отличаются друг от друга по морфологии, биохимическим и функциональным свойствам. В основе этих различий лежит активное функционирование в разных клетках неодинаковых частей генома. Большая часть генома находится в клетках организма в неактивном, репрессированном, состоянии, и только 7—10% генов активны, т. е. транскрибируются. Спектр функционирующих генов зависит от тканевой принадлежности клетки, от  периода ее жизненного цикла и стадии индивидуального развития организма. Основная масса генов, активно функционирующих в большинстве клеток организма на протяжении онтогенеза,— это гены «домашнего хозяйства», которые обеспечивают синтез белков общего назначения (белки рибосом, гистоны, тубулины и т. д.), тРНК и рРНК. Транскрибирование этих генов обеспечивается соединением РНК-полимеразы с их промоторами и, видимо, не подчиняется каким-либо другим регулирующим воздействиям. Такие гены называют конститутивными. Другая группа генов, детерминирующих синтез специфических продуктов, в своем функционировании зависит от различных регулирующих факторов, ее называют регулируемымигенами. Они функционируют только в определенных клетках и тканях и называются генами «роскоши». Изменение условий может привести к активации «молчащих» генов и репрессии активных. Дифференцированная экспрессия одного генома у млекопитающих обусловливает развитие огромного множества типов клеток. 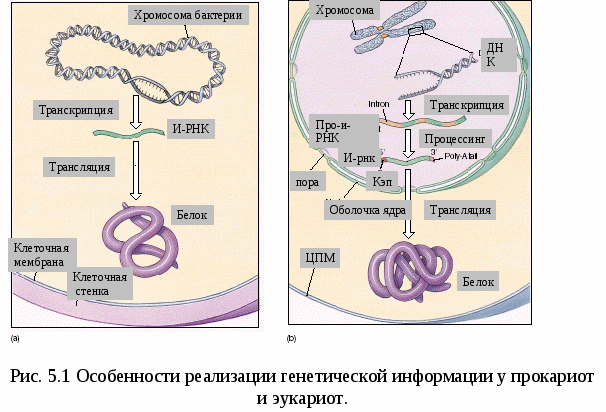 Глава 5 Особенности структурной и функциональной организации генов про- и эукариот. Регуляция экспрессии генов у про- и эукариот. Реализация генетической информации. Биосинтез белка Наследственность и изменчивость - важнейшие свойства любой живой системы, обеспечиваются функционированием особого материального субстрата. Как же он устроен? Первые предположения об организации наследственного материала высказал основоположник генетики Г.Мендель. В своей работе “Опыты над растительными гибридами ” на основании результатов проведенных экспериментов на горохе он пришел к заключению, что наследственный материал дискретен, т.е. представлен отдельными наследственными задатками, отвечающими за развитие определенных признаков. Понятия «ген» в то время еще не существовало. Только в 1909 году В. Иогансен назвал «наследственные задатки» Менделя генами. Что же такое ген? Следует сразу оговориться, что до сих пор не существует достаточно полного и всеобъемлющего определения понятия ген, оно эволюционировало вместе с познанием его структуры и функции. Долгое время ген рассматривали как минимальную часть наследственного материала, обеспечивающую развитие определенного признака, что выражалось формулой «1 ген—1 признак». Это понятие классической генетики ХХ века. Между тем, какова структура гена, как он функционирует оставалось неясным. В 1945 г. Бидл и Татум установили, что гены отвечают за образование определенных ферментов, которые затем оказывают влияние на развитие тех или иных признаков. Эту гипотезу можно выразить формулой «1 ген-1 фермент» или «1 ген-1 белок». Однако позднее было показано, что многие белки имеют четвертичную структуру, в образовании которой принимают участие разные пептидные цепи. Например, гемоглобин взрослого человека включает 4 глобиновые цепи - 2 альфа и 2 бетта, которые кодируются совершенно разными генами. Поэтому формула гена была преобразована: «1 ген – 1 полипептид». Но в дальнейшем открытие особенностей структурной организации генов показало, что одна и та же последовательность (т.е. ген) может обеспечивать синтез нескольких разных полипептидных цепей с разными функциями. Так, у человека примерно 100.000 белков и всего 30.000 генов, т.е. большинство генов несут информацию о нескольких полипептидах. Стало понятно, что нельзя говорить о гене как об участке генома, кодирующем только 1 полипептидную цепь. Кроме того, в гене Оперон 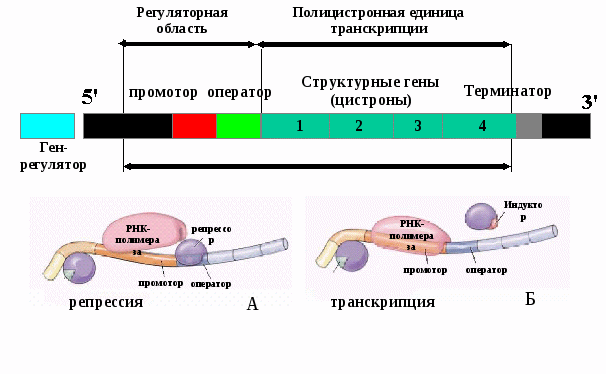 Рис. 5.2. Схема регуляции экспрессии генов у прокариот (А - Белок-репрессор блокирует оператор, Б - Индуктор связывает белок-репрессор. Оператор освобождается. РНК-полимераза осуществляет транскрипцию). зашифрована информация не только о каком-то белке, но и об РНК (н-р, тРНК). Таким образом, в настоящее время определение гена следующее: Ген – это фрагмент ДНК, ассоциированный с регуляторными элементами и соответствующий одной единице транскрипции, которая определяет возможность синтеза полипептидной цепи или молекулы РНК. Реализация наследственной информации, заключенной в генотипе организма,— это сложный процесс, который требует тонкой регуляции для того, чтобы в клетках разной тканевой принадлежности в определенное время в процессе развития организма обеспечить синтез специфических белков в необходимом количестве (рис. 5.1). Рассмотрим основные закономерности экспрессии (функционирования) генов. Экспрессия генов у прокариот Рассмотрим классическую схему работы (экспрессии) генов прокариот по принципу обратной связи или негативного контроля (репрессии) на примере лактозного оперона у бактерий (кишечной палочки). Эта схема была предложена французскими учеными Жакобом и Моно в 1961 г. За эту работу, признанную классической, они были удостоены Нобелевской премии. В состав оперона у прокариот входят промотор, ген-оператор (включает или выключает работу структурных генов), группа регуляторных и структурных генов и терминатор (рис. 5.2.). На некотором расстоянии от оперона находится генрегулятор, который синтезирует белки 2х типов. Белок-репрессор блокирует оператор, оперон не работает, белок не синтезируется (рис. 5.2 А). Если в клетку поступает индуктор, то он связывает белокрепрессор. Оператор освобождается, происходит считывание информации с ДНК на мРНК, запускается биосинтез белка (рис. 5.2 Б). Накопление белка (фермента) избыточном количестве приводит к связыванию и разрушению индуктора. Белокрепрессор освобождается, оператор блокируется и работа оперона останавливается. |