Лекции биохимия. Лекция Предмет и методы биохимии. Строение, свойства и биологические функции углеводов и липидов
 Скачать 2.08 Mb. Скачать 2.08 Mb.
|

СВОЙСТВА АМИНОКИСЛОТЧистые препараты аминокислот - белые кристаллические вещества, имеющие высокие температуры плавления (свыше 200˚С). Многие из них хорошо растворяются в воде, за исключением кислот с гидрофобными радикалами. В водном растворе карбоксильные группы аминокислот подвергаются диссоциации с образованием катионов водорода, а азот аминогруппы довольно активно присоединяет катионы водорода, превращаясь в заряженную группировку, в результате чего образуется биполярное соединение: 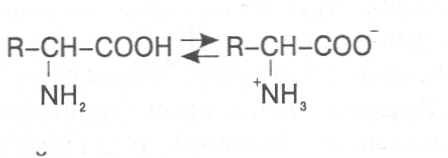 В сильно кислой среде диссоциация карбоксильных групп подавляется высокой концентрацией ионов водорода и в таких условиях заряд молекулы будет положительным, так как он определяется зарядом атома азота, присоединившего протон: 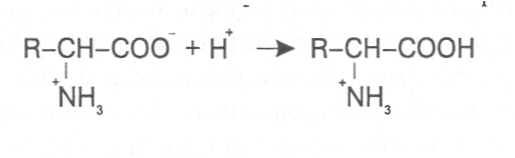 При понижении концентрации ионов водорода (увеличение рН) степень диссоциации карбоксильных rpyпп yвeличивaeтcя, а взаимодействие ионов водорода с азотом аминогруппы ослабляется, вследствие чего при определенном значении рН большая часть карбоксильных групп будет диссоциировать и молекула аминокислоты приобретает отрицательный заряд. В щелочной среде под воздействием высокой концентрации гидроксид-ионов происходит связывание катионов водорода, присоединенных к аминогруппе, с образованием воды, в результате чего заряд молекулы будет определяться только зарядом карбоксильной группы: 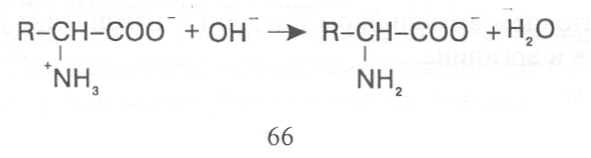 При определенном значении рН среды положительный заряд аминогрупп полностью уравновешивается отрицательным зарядом карбоксильных групп и молекулы аминокислоты становятся элекrpонейтральными. Такое состояние молекулы принято называть изоэлектрической точкой. Если рН среды ниже рН изоэлектрической точки молекулы аминокислоты, то она заряжена положительно за счет заряда аминогруппы и ведет себя в водном растворе как катион. При рН среды выше рН изоэлектрической точки молекула аминокислоты заряжена отрицательно за счет диссоциации карбоксильной группы и ведет себя в водном растворе как анион. Опытным путём определено, что изоэлектрические точки моноаминомонокарбоновых кислот находятся в слабокислой среде(рН 5-6,5), так как кислотные свойства у них выражены несколько сильнее, чем основные. У моноаминодикарбоновых кислот кислотные свойства выражены значительно сильнее, поэтому их изоэлектрические точки находятся в кислой среде (рН 3-3,2). У диаминомонокарбоновых кислот сильнее выражены основные свойства, в связи с чем их изоэлектрические точки смещены в щелочную сторону (рН 9,7-10,8). В связи с тем, что в молекулах аминокислот имеются как кислотные, так и основные группировки, они проявляют амфотерные свойства и могут взаимодействовать и с кислотами, и с основаниями, образуя соответствую-щие соли: 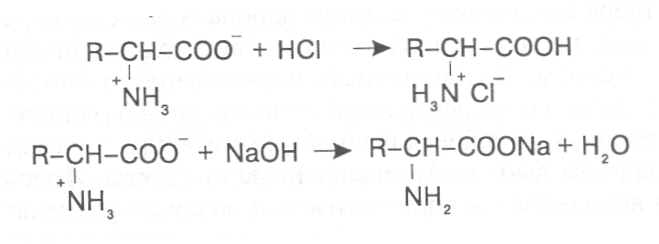 Своими карбоксильными и аминными группировками аминокислоты могут вступать и в другие химические реакции, с помощью которых можно проводить качественный и количественный анализ этих соединений. При взаимодействии аминокислот со спиртами образуются сложные эфиры, которые подвергаются вакуумной перегонке при разных физических параметрах. Благодаря этому можно разделять смеси аминокислот путём получения и фракционной перегонки их эфиров. Аминокислоты довольно легко вступают в реакции с редуцирующими сахарами, образуя при этом темноокрашенные продукты–меланоидины. На первом этапе взаимодействия аминокислоты и моносахарида образуются продукты их разложения: из сахаров – фурфурол или оксиметилфурфурол, а из аминокислоты – альдегид, углекислый газ и аммиак. На следующем этапе фурфурол или оксиметилфурфурол, вступая в соединение с аминокислотами, образуют меланоидины. Интенсивность реакции усиливается при повышенной температуре, которая создается при термической обработке рacтитeльной продукции или её перерaботке (сушка овощей, фруктов и продуктов из картофеля, выпечка хлеба, получение макарон и кондитерских изделий). Аналогичные явления наблюдаются при самосогревании зерна. Промежуточные продукты меланоидинообразования – альдегиды создают специфический запах, характерный для тех или иных пищевых продуктов. Фурфурол имеет запах яблок, оксиметилфурфурол – запах мёда, изовалериановый альдегид, образуемый из аминокислоты лейцина, – запах ржаного хлеба. Конечные продукты реакций взаимодействия аминокислот с редуцирующими сахарами – меланоидины вызывают потемнение растительных продуктов, что ухудшает их товарные свойства. В процессе хлебопечения реакции образования меланоидинов влияют на формирование цвета, вкуса и аромата ржаного и пшеничного хлеба. В результате ферментативного окисления ароматических аминокислот тирозина и фенилаланина также образуются темноокрашенные продукты - меланины, вызывающие потемнение на воздухе очищенных клубней картофеля, измельченных корнеплодов, овощей и фрyкroв. Эти вещества совместно с меланоидинами определяют тёмный цвет ржаного хлеба и вызывают потемнение макарон при сушке. Меланины синтезируются в ходе крнденсации продуктов окисления тирозина и фенилаланина, возникающих под действием фермента тирозиназы (см. стр. …). 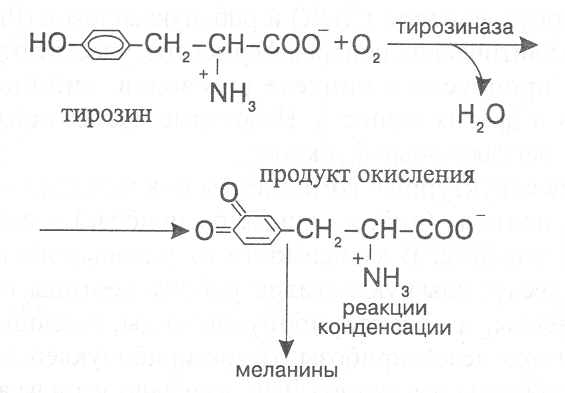 В целях предотвращения потемнения растительных продуктов при хранении, сушке и переработке применяются технологии выращивания сельскохозяйственных культур, не допускающие накопления в товарной продукции редуцирующих сахаров и свободных аминокислот. 4.2. Строение, свойства и биологические функции нуклеотидов. Наряду с аминокислотами важнейшей группой азотистых веществ являются нуклеотиды. Их биологическое значение для жизнедеятельности организмов определяется тем, что они используются для построения молекул нуклеиновых кислот - дезоксирибонуклеиновой (ДНК) и рибонук-леиновой (РНК), входят в состав каталитических центров ферментов, участвуют в биоэнергетических процессах и синтезе углеводов, липидов, белков, алкалоидов и других веществ. Некоторые нуклеотиды способны выполнять регуляторные функции. Главные структурные компоненты нуклеотидов–азотистые основания, пентозы (рибоза или дезоксирибоза) и остаток ортофосфорной кислоты. В зависимости от углеводного компонента различают две группы нук-леотидов: рибонуклеотиды, содержащие остаток рибозы, и дезоксирибо-нуклеотиды, имеющие в своем составе остаток дезоксирибозы. Дезоксирибонуклеотиды используются организмами для синтеза ДНК, а рибонуклетиды входят в состав РНК, ферментов и макроэргических нук-леозидполифосфатов. Рибоза и дезоксирибоза в составе нуклеотидов находятся в b-D-фура-нозной форме:  Нуклеотиды образуются из двух типов азотистых оснований – произ-водных пиримидина и пурина. Свойства оснований они проявляют в водном растворе при взаимодействии с молекулами воды. Из пиримидиновых осно-ваний наиболее важное значение имеют урацил, тимин и цитозин как основные структурные единицы нуклеотидов, образующих нуклеиновые кислоты. Кроме них, известны и другие основания - 5-метилцитозин, псевдоурацил, 5-оксиметилцитозин и др. 5-Метилцитозин и 5-оксиметилцитозин в небольшом количестве могут 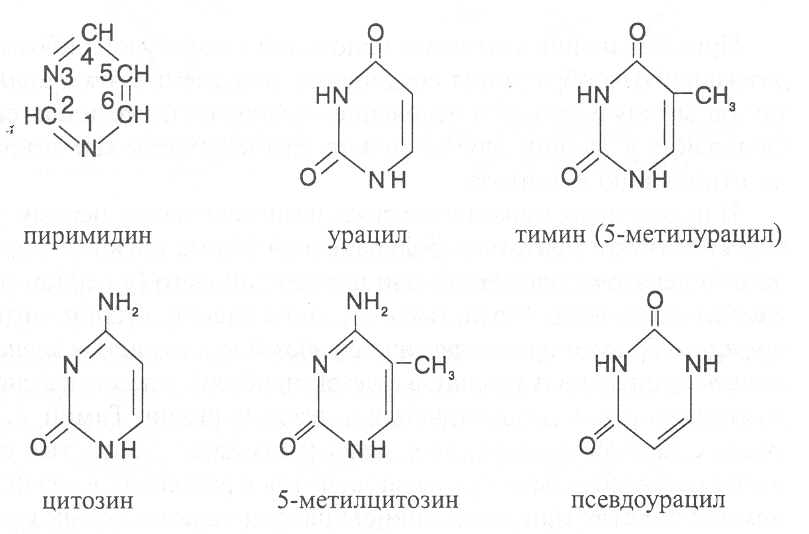 содержаться в составе нуклеотидов ДНК и РНК, псевдоурацил - в транспортной РНК. Из пуриновых оснований наибольшее значение имеют аденин и гуанин, так как они используются для синтеза нуклеиновых кислот. В составе нуклеиновых кислот в небольшом количестве обнаружены также и другие основания, которые образуются в результате химической модификации аденина и гуанина: 7-метилгуанин, 2-метиладенин, N-диметилгуанин и др. Важными промежуточными метаболитами являются гипоксантин, ксантин, аллантоин. В некоторых растениях они могут накапливаться в свободном состоянии. Все азотистые основания интенсивно поглощают ультрафиолетовый свет при длинах волн 200-280нм. При соединении азотистых оснований с молекулой рибозы или дезоксирибозы образуются соединения, называемые нуклеозидами, так как между пентозой и основанием возникает гликозидная связь. Основания в данном случае можно рассматривать как агликоны по отношению к пентозе. 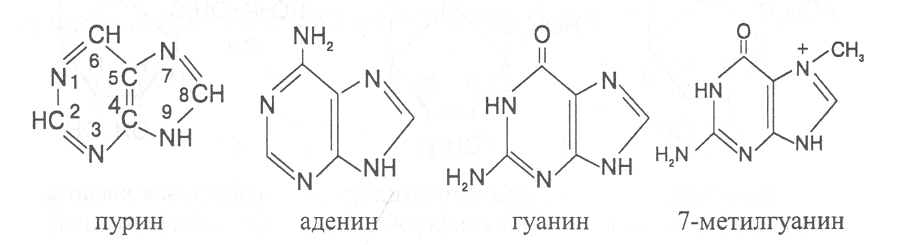 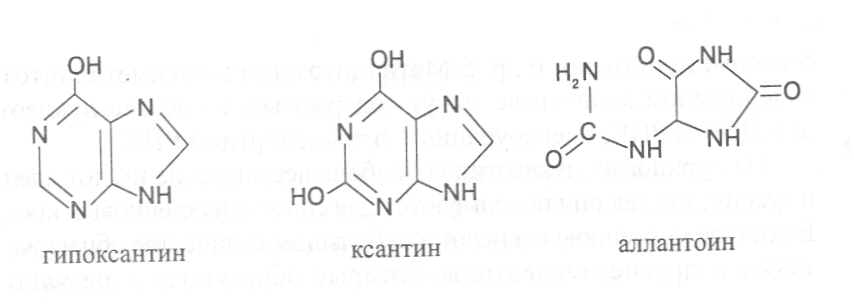 В нуклеозидах гликозидная связь возникает между первым углеродным атомом пентозы в b-фуранозной форме и азотом пуринового (в девятом положении) или пиримидинового (в первом положении) основания. Азотистые основания аденин, гуанин, цитозин и урацил образуют при со-единении с рибозой нуклеозиды - аденозин, гуанозин, цитидин и уридин, 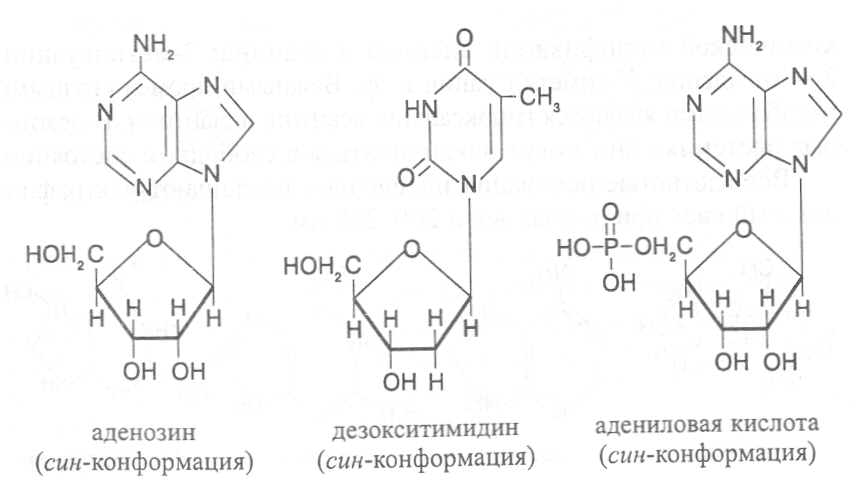 а с дезоксирибозой – дезоксиаденозин, дезоксигуанозин, дезоксицитидин, дезоксиуридин. Тимин, соединяясь с дезоксирибозой, даёт дезоксити-мидин. 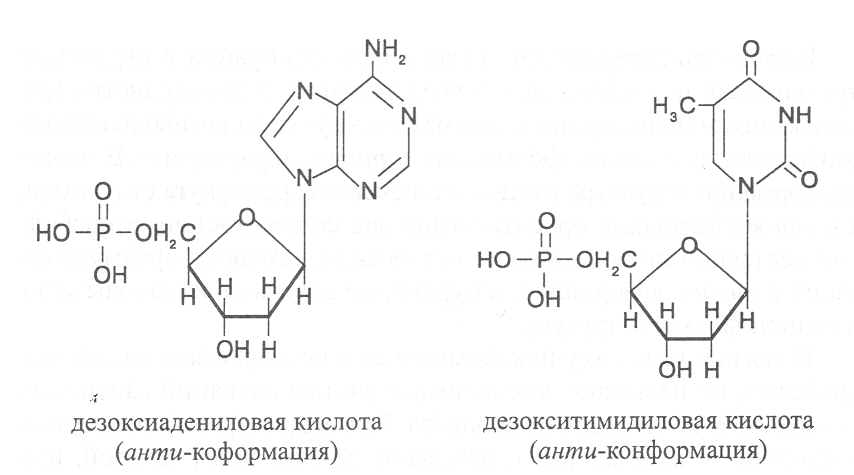 Азотистые основания и нуклеозиды могут накапливаться в растениях в значительном количестве при интенсивном распаде нуклеиновых кислот. Фосфорнокислые эфиры нуклеозидов называют нуклеотидами. В составе нуклеотидов остатки ортофосфорной кислоты могут присоединяться к пятому или третьему атомам углерода рибозы или дезоксирибозы, а у некоторых рибонуклеотидов ещё и ко второму атому углерода рибозы. У свободных нуклеотидов фосфатная группа обычно находится у пятого углеродного атома рибозы или дезоксирибозы. В нейтральной среде остатки ортофосфорной кислоты в молекулах нуклеотидов сильно диссоциированы, вследствие чего могут присоединять катионы, поэтому при химическом выделении нуклеотиды кристаллизуются в виде солей. Изучение пространственной структуры азотистых оснований методом рентгеноструктурного анализа показывает, что все они имеют почти плоскую конформацию. У них довольно легко происходит перегруппировка двойных связей, которая сопровождается таутомерными превращениями. Например, гуанин может существовать в виде двух таутомерных форм: 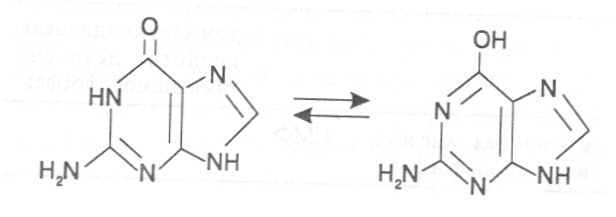 Плоскость гетероциклического ядра основания в структуре нуклеозидов и нуклеотидов может занимать в пространстве два положения по отношению к пентозе, образуя две противоположные конформации - син-конформацию и анти-конформацию. В анти-конформации структура азотистого основания развернута от пентозы, а в син-конформации ориентирована над её плоскостью. В свободном состоянии пиримидиновые нуклеотиды находятся преимущественно в анти-конформации, а пуриновые довольно легко переходят из одной формы в другую. В связи с тем, что у нуклеотидов сильно выражены кислотные свойства, их называют кислотами с учетом названий азотистых оснований и углеводного компонента. Так, например, рибонуклеотид, имеющий остаток аденина, называют адениловой кислотой, или аденозинмонофосфатом (АМФ). Дезоксирибонуклеотид, образованный из тимина, называют дезокситимидиловой кислотой, или дезокситимидинмонофосфатом (дТМФ). Названия других нуклеотидов представлены в таблице 2. В растениях найдены циклические формы нуклеотидов – адено-зинмонофосфата и гуанозинмонофосфата, которые по-видимому выполняют регуляторные функции. Строение циклического АМФ можно представить слудующей формулой: 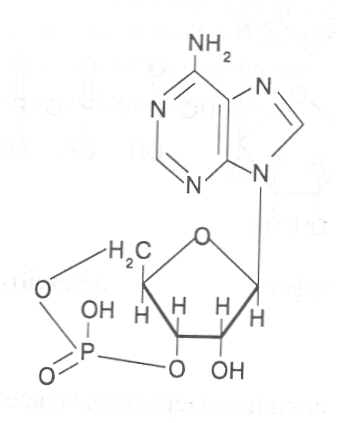 2. Названия важнейших нуклеотидов.
Путём фосфорилирования нуклеотиды превращаются в более активные формы - нуклезиддифосфаты и нуклеозидтрифосфаты, в виде которых, как мы увидим далее, они участвуют в различных биохимических процессах – синтезе нуклеиновых кислот, сложных углеводов и липидов, коферментов и витаминов, запасании и переносе энергии. Из адениловой кислоты образуются аденозинди- и аденозинтрифосфат (АДФ и АТФ), из гуаниловой кислоты - гуанозинди- и гуанозинтрифосфат (ГДФ и ГТФ), из цитидиловой кислоты - цитидинди- и цитидинтрифосфат (ЦДФ и ЦТФ), из уридиловой кислоты - уридинди- и уридинтрифосфат (УДФ и УТФ), из инозиновой кислоты – инозинди- и инозинтрифосфат (ИДФ и ИТФ). Из дезоксирибонуклеотидов также синтезируются соответствующие ди- и трифосфаты: дезоксиаденозинди- и дезоксиаденозинтрифосфат (дАДФ и дАТФ), дезоксигуанозинди- и дезоксигуанозинтрифосфат (дГДФ и дГТФ), дезоксицитидинди- и дезоксицитидинтрифосфат (дЦДФ и дЦТФ), де-зокситимидинди- и дезокситимидинтрифосфат (дТДФ и дТТФ). Строение ди- и трифосфатов нуклеозидов можно представить следующими формулами: 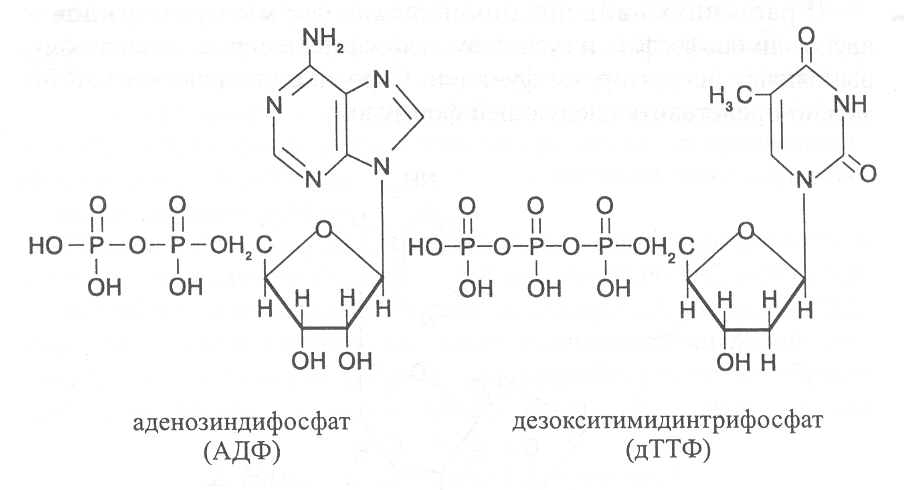 Свободные нуклеотиды содержатся во всех растительных клетках. Они возникают или в процессе активного синтеза азотистых веществ, или в результате распада нуклеиновых кислот, который может интенсивно происходить при неблагоприятных условиях произрастания, а также при хранении и переработке растительных продуктов. Так, например, при разваривании клубней картофеля образующиеся в результате распада нуклеиновых кислот свободные нуклеотиды оказывают влияние на формирование вкусовых качеств варёного картофеля. По интенсивности образования различных фракций нуклеотидов можно судить о направленности биохимических процессов в организме. Если в клетках и тканях организма повышается содержание трифосфатпроизводных нуклеотидов, то это свидетельствует об активизации его жизнедеятельности, тогда как накопление монофосфатов наблюдается при усилении процессов распада веществ, характерных для стареющего организма. 4.3. Строение, свойства и биологические функции белков. Белки - неотъемлемые компоненты любой живой клетки, которые обеспечивают и поддерживают ее жизнедеятельность. Молекулы белков представляют собой биополимеры, построенные в основном из амино-кислот. Кроме аминокислот в состав белковых молекул могут входить другие органические и неорганические компоненты. В белках содержится 50-55% углерода, 20-24% кислорода, 7% водорода, 0,5-3% серы; в состав некоторых белков могут также входить фосфор и различные металлы. Огромное структурное разнообразие белков и широкий диапазон изменения их физико-химических свойств позволяют этим биополимерам выполнять разнообразные и жизненно важные функции в живом организме. В каждой растительной клетке одновременно функционируют несколько тысяч различных белков. Все биохимические реакции в клетке происходят с участием каталитических белков - ферментов. Структурная основа биологических мембран цитоплазмы и внутриклеточных органоидов также построена с участием белков. Защитную функцию выполняют белковые антитела и стрессовые белки, образующиеся под воздействием стрессовых факторов. Важную роль выполняют в растительных клетках регуляторные и транспортные белки, способные обратимо изменять свою конформацию и таким образом активно участвовать в поддерживании жизнедеятельности растения как саморегулирующейся системы. В семенах и других органах растений откладываются запасные бел-ки, которые в значительной степени определяют питательную, кормовую и технологическую ценность растительной продукции. Много белков на-капливается в зерне зернобобовых культур - 20-30%, в сое и люпине - 30-40%, в семенах масличных культур - 15-30%. Содержание белков в другой растительной продукции составляет, %: зерновки злаковых растений – 9-18; кукуруза и рис – 6-10; клубни картофеля – 1,5-2; корнеплоды – 1-1,5; овощи, плоды и ягоды – 0,5-2; цветная капуста – 2-3; брюссельская капуста и чеснок –6-8; вегетативная масса мятликовых трав – 5-15, бобовых трав – 15-25 (последние два показателя даны в расчёте на сухую массу). |