Лекции биохимия. Лекция Предмет и методы биохимии. Строение, свойства и биологические функции углеводов и липидов
 Скачать 2.08 Mb. Скачать 2.08 Mb.
|

СН3(СН2)5СН(ОН)СН2СН=СН(СН2)7СООНВ маслах из семян растений семейства капустные (рапс, горчица, рыжик) содержится мононенасыщенная эруковая кислота: СН3(СН2)7СН=СН(СН2)11СООНВедётся направленная селекция этих растений с целью понижения в масле содержания эруковой кислоты. Каждый вид растений имеет совершенно определённый набор жирных кислот в составе масла. Близкие между собой генотипы почти не отличаются по составу масла. В то же время удельное соотношение жирных кислот в масле одного и того же вида растений существенно изменяется в зависимости от фазы развития и условий внешней среды. Жирные кислоты, входящие в состав масла, образуют с глицерином смесь эфиров, которая включает разные по кислотному составу ацилглицерины. Они могут быть однокислотные, если содержат остатки одной кислоты, или разнокислотные с разными комбинациями кислотных радикалов. Так, например, в оливковом масле, в котором преобладает олеиновая кислота, довольно много содержится триолеина, а в касторовом масле с участием рицинолевой кислоты образуется однокислотный ацилглицерин - тририцинолеин. Во многих растительных маслах содер-жатся ацилглицерины, образующиеся из пальмитиновой, олеиновой и линолевой кислот - пальмитинодиолеин и пальмитиноолеинолинолеин. В результате различных комбинаций с участием нескольких жирных кислот формируется довольно большое разнообразие триацилглицеринов, характерных для каждого вида растений. 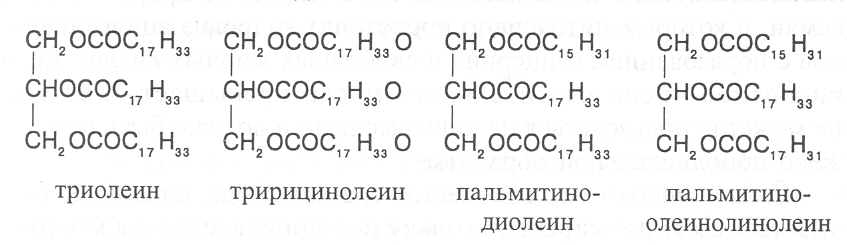 В нерафинированном растительном масле, кроме ацилглицеринов, в растворенном состоянии находятся свободные жирные кислоты и фос-фолипиды (1-2%), стероидные липиды (0,1-1,5%), жирорастворимые ви-тамины и пигменты. Вследствие того, что в большинстве растительных масел растворены каротиноиды, они окрашены в желтый цвет. Для получения кулинарных жиров, а также производства мыла разработаны промышленные способы превращения растительных масел в твердые жиры. С этой целью проводится гидрогенизация жидких растительных жиров в присутствии катализаторов, при которой в результате присоединения водорода происходит превращение в составе ацилглицеринов ненасыщенных кислот в насыщенные, вследствие чего жир приобретает твердую консистенцию. Для характеристики свойств жира применяют показатели, называемые числами жиров; наиболее важное значение имеют кислотное число, йодное число и число омыления. При указании значений этих чисел размерность обычно не записывается. Кислотное число выражается количеством миллиграммов гидроксида калия, необходимого для нейтрализации свободных жирных кислот, содержащихся в 1 г жира. Оно характеризует содержание свободных жирных кислот в жире. В масле из созревших семян концентрация свободных кислот минимальная, поэтому кислотное число такого масла имеет низкое значение (не более 1-2). Однако масло из недозревших семян содержит много свободных жирных кислот и его кислотное число увеличивается. Ещё большее увеличение кислотного числа наблюдается в маслах из проростающих семян, в которых интенсивно происходит гидролиз ацилглицеринов с образованием глицерина и свободных жирных кислот. Если кислотное число растительного масла превышает 5, то оно не может использоваться на пищевые цели и должно быть подвержено дополнительной обработке. Йодное число - это количество граммов йода, способное связываться со 100 г жира. Поскольку йод присоединяется к жирам при разрыве двойных связей в радикалах ненасыщенных жирных кислот, этот показатель характеризует степень непредельности ацилглицеринов. Чем больше двойных связей в кислотных остатках, тем выше йодное число жира. Йодные числа животных жиров, содержащих остатки насыщенных кислот, имеют низкие значения (30-70). Растительные жиры, образованные главным образом из ненасыщенных кислот, отличаются более высокими йодными числами (80-180). Число омыления - количество миллиграммов гидроксида калия, необ- ходимое для нейтрализации свободных и связанных в составе ацилглицеринов жирных кислот, содержащихся в 1 г жира. Оно характеризует среднюю величину молекулярной массы жирных кислот и образующихся из них ацилглицеринов, входящих в состав жира. Ацилглицерины, включающие радикалы низкомолекулярных жирных кислот, имеют более высокое число омыления. Число омыления животных жиров и жиров растений умеренных широт обычно находится в пределах 170-200, а жиров тропических растений - 200-250. ПРОГОРКАНИЕ ЖИРОВ. При длительном хранении жиры под воздействием ряда факторов подвергаются прогорканию, вследствие чего они приобретают неприятный вкус и запах. Одной из причин прогоркания растительных масел и содержащих их продуктов является действие ферментов - липаз и липоксигеназ. Под действием липаз происходит гидролиз сложноэфирных связей ацилглицеринов с образованием глицерина и свободных жирных кислот. При этом некоторые жирные кислоты имеют неприятный вкус и запах. Фермент липоксигеназа катализирует окисление свободных жирных кислот, превращая их в гидроперекиси: 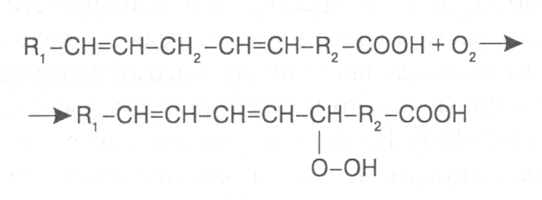 Гидроперекиси, в свою очередь, как сильные окислители, подвергают жирные кислоты дальнейшему окислению с образованием альдегидов и кетонов, обладающих неприятным вкусом и запахом, характерным для процесса прогоркания жиров. Может также происходить неферментативное прогоркание жиров, связанное с воздействием на них влаги, света и кислорода воздуха. На первом этапе жир подвергается гидролизу, а затем ненасыщенные жирные кислоты окисляются кислородом воздуха с образованием перекисей: 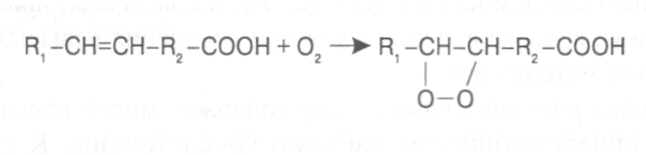 Затем перекиси подвергаются разложению, образуя низкомолекулярные кислоты, а также альдегиды и кетоны с неприятным запахом. Процесс прогоркания жиров могут также инициировать продукты жизнедеятельности микроорганизмов. Для замедления окислительного прогоркания растительных масел к ним добавляют вещества - антиокислители, предохраняющие их от окисления. В растительном масле содержится также природный антиокислитель - токоферол (витамин E). ВЫСЫХАНИЕ МАСЕЛ. Под действием кислорода происходит также высыхание жиров, имеющих в своем составе радикалы полиненасыщенных жирных кислот. В процессе окисления этих кислот происходит их деградация по двойным связям с образованием углекислого газа, воды и летучих альдегидов. Одновременно происходит полимеризация масел. Окисляющийся жир постепенно густеет на воздухе и образует эластичную пленку, которая не растворяется в органических растворителях и устойчива к различным внешним воздействиям. Эти свойства растительных жиров используются для приготовления олифы, лаков и красок. По способности к высыханию и в зависимости от состава жирных кислот растительные масла разделяют на четыре группы. Хорошо высыхающие масла имеют высокое содержание полиненасы- щенных жирных кислот и характеризуются высокими йодными числами (140-180). Их используют как сырье для лакокрасочной промышленности. К таким маслам относятся льняное, конопляное, тунговое, перилловое. Образующаяся при их высыхании пленка не растрескивается под воздействием влаги и солнечных лучей и сохраняет эластичность. Слабо высыхающие масла имеют йодные числа в пределах 100-130. Образующаяся при их высыхании пленка растрескивается на воздухе. К этой группе относятся пищевые масла - подсолнечное, соевое, хлопковое, оливковое, горчичное, рапсовое, кукурузное и др. Невысыхающие масла - касторовое, арахисовое. Они имеют низкие йодные числа (80-100) и содержат в своем составе специфические жирные кислоты (рицинолевую, арахиновую), определяющие их невысыхаемость на воздухе. Эти масла находят применение в медицине, а также в технике для приготовления невысыхающих смазочных материалов. Твёрдые растительные масла содержат много насыщенных кислот, определяющих их твёрдую консистенцию. К твёрдым относятся масла многих тропических растений - пальмовое, кокосовое, масло бобов какао и др. Селекционерами совместно с генетиками и биохимиками проводятся работы по созданию новых сортов масличных культур с повышенным накоплением масла и заданным составом жирных кислот. Жиры обладают высокой энергетической ценностью и поэтому ис-пользуются живыми организмами в качестве запасных веществ. При их окислении выделяется значительно больше энергии в расчёте на единицу массы, чем при окислении углеводов или белков. Определено, что энергетическая ценность 1 г жира составляет в среднем 39 кДж энергии, углеводов - 17-20 кДж, белков - 22-24 кДж. В результате изучения химического состава растений выяснено, что более чем у 90% растительных видов основными запасными веществами семян являются жиры. Следует также учитывать, что при окислении жиров по сравнению с белками и углеводами выделяется значительно больше и воды, которая имеет важное биологическое значение для организмов. Для оценки накопления жиров в растительных продуктах очень часто используют показатель - сырой жир, который выражает общее содержание липидов, переходящих при экстракции в растворитель – диэтиловый эфир. В сыром жире основной липидный компонент – это собственно жир, однако в определенном количестве в экстракт переходят и другие фракции липидов, которые находятся в несвязанном состоянии. 3.2. Строение, свойства и биологические функции фосфолипидов. Фосфолипиды - белые воскообразные вещества, хорошо растворимые в органических растворителях - эфире, бензоле, хлороформе. На воздухе они быстро окисляются и темнеют. От жиров фосфолипиды отличаются наличием в их молекулах фосфатной группы, к которой сложноэфирной связью присоединяется азотистое или другое соединение. Строение фосфолипидов можно представить следующей формулой: 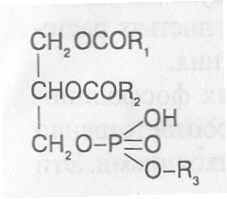 В этой формуле R₁ и R₂ – радикалы жирных кислот, а R₃ – остаток азотистого или другого соединения. В состав фосфолипидов чаще всего входят пальмитиновая, стеариновая, олеиновая и линолевая кислоты, причём ненасыщенная кислота связана со вторым углеродным атомом глицеринового остатка. Молекулы фосфолипидов обладают заметно выраженной полярностью. Углеводородные радикалы жирных кислот представляют гидрофобную часть молекулы, а остатки азотистых соединений, глицерина, спирта инозита проявляют гидрофильные свойства, вследствие чего в водном растворе и на границе двух фаз они приобретают определённую ориентацию. Благодаря этим свойствам фосфолипиды играют важную роль в формировании структуры клеточных мембран. В составе мембран они находятся в соединении с белками в виде липопротеидов и могут участвовать в регуляторных процессах. Как поверхностноактивные вещества фосфо-липиды используются в качестве эмульгаторов при изготовлении кондитерских изделий. Они улучшают хлебопекарные свойства пшеничной муки. В семенах растений фосфолипиды откладываются в качестве запасных веществ, повышая таким образом их пищевую и кормовую ценность. В зерновках злаковых растений содержание фосфолипидов составляет 0,2-0,6%, а в семенах масличных и бобовых культур - 1-2%, в зародышах различных семян - 1,5-3%. Простейшими фосфолипидами следует считать фосфатидные кислоты, которые представляют собой диацилглицерины, соединённые слож-ноэфирной связью с остатком ортофосфорной кислоты: 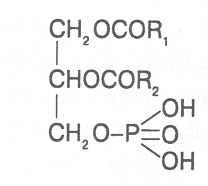 Фосфатидные кислоты содержатся в растениях в небольших количествах, так как являются промежуточными продуктами липидного обмена. Они найдены в зародышах семян и в листьях растений в виде солей с катионами кальция, калия и магния. Значительно больше синтезируется в растениях фосфолипидов, имеющих остатки этаноламина и холина, их соответственно называют фосфатидилэтаноламинами и фосфатидилхолинами. Они входят в состав клеточных мембран и откладываются в семенах в качестве запасных веществ. Фосфатидилэтаноламины и фосфатидилхолины образуют смеси близких по свойствам липидов, различающихся остатками жирных кислот. 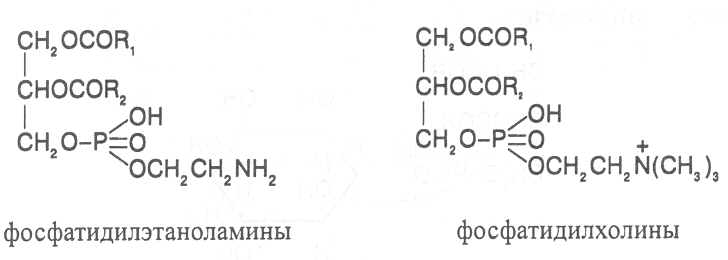 В составе митохондриальных и хлоропластных мембран содержатся фосфолипиды, у которых к фосфатидной кислоте сложноэфирной связью присоединяется остаток аминокислоты серина или глицерина, их называют соответственно фосфатидилсеринами и фосфатидилглицеринами. 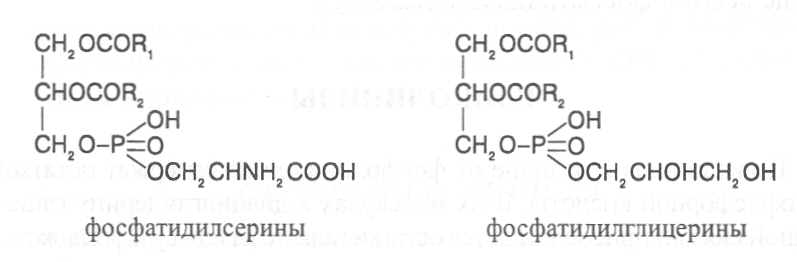 Фосфатидилглицерины составляют почти половину от всех липидов хлоропластных мембран и в их молекулах находятся остатки транс-изомера ненасыщенной гексадеценовой кислоты: 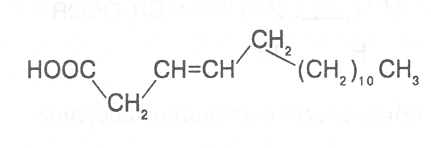 В клеточных мембранах многих растений и некоторых водорослей обнаружены фосфолипиды, у которых с фосфатидной кислотой связаны остатки моносахаридов (глюкозы, галактозы, арабинозы), а также одного из изомеров циклического спирта инозита – миоинозита. 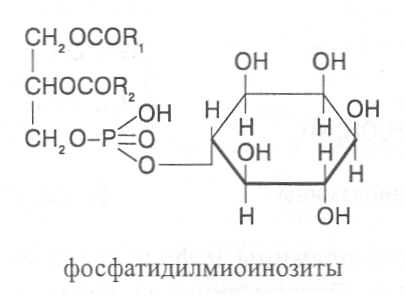 С участием миоинозита и указанных моносахаридов могут синтезироваться более сложные фосфолипиды, у которых между миоинозитом и одним из моносахаридов образуется гликозидная связь, а миоинозит полученного гликозида через остаток ортофосфорной кислоты присоединяется к какому-либо фосфолипиду (чаще всего к фосфати-дилэтаноламину). 3.3. Строение, свойства и биологические функции гликолипидов. Гликолипиды в отличие от фосфолипидов не содержат остатков ортофосфорной кислоты. В их молекулах к диацилглицерину гликозидной связью присоединяются остатки галактозы или сульфоглюкозы. 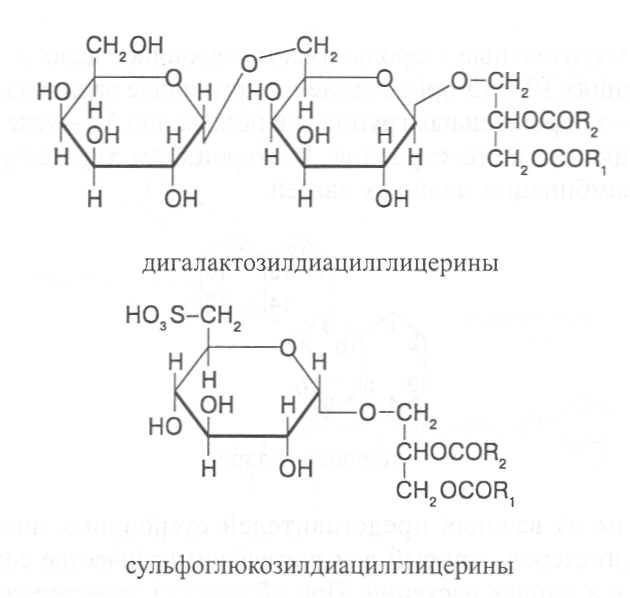 Такие липиды в большом количестве содержатся в мембранах хлоропластов растений и некоторых водорослей. В галактозилдиацил-глицеринах основной ненасыщенной жирной кислотой является линоленовая кислота. 3.4. Строение, свойства и биологические функции стероидных липидов. Стероидные липиды, или стеролы - это полициклические спирты, производные циклопентанопергидрофенантрена, которые присутствуют в клетках растений в свободном виде или в составе гликозидов. Образуя с белками сложные комплексы, они участвуют в построении клеточных мембран. Стероиды хорошо растворяются в органических и жировых растворителях, поэтому содержатся в растительных маслах. В семенах растений содержание стероидных липидов колеблется в пределах 0,05-1,5%, в вегетативных частях - 0,05-0,2% (в расчёте на сухую массу). Особенно много стеролов в дрожжах (свыше 2%). Все растительные стеролы имеют стероидное ядро, к которому в положениях 10 и 13 присоединены метильные радикалы, в положении 3 - гидроксильная группа, а в положении 17 - углеводородный радикал разного строения. В стероидном ядре встречаются разные комбинации двойных связей. 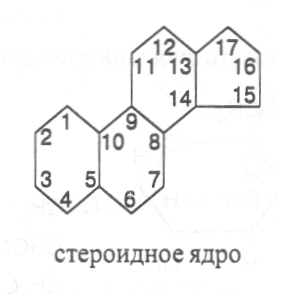 Одним из важных представителей стероидных липидов является эргостерол, который в значительном количестве содержится в листьях и плодах растений. При облучении эргостерола ультрафиолетовыми лучами его молекула подвергается структурной модификации, превращаясь в витамин D2 (эргокальциферол). Препараты этого витамина получают в большом количестве из эргостерола, образующегося в клетках дрожжей. В семенах растений в довольно большом количестве содержатся стероидные липиды, не способные превращаться в витамины. К ним от-носятся b-ситостерол, стигмастерол, спинастерол, кампестерол, холестерол и некоторые другие. Все они являются полициклическими спиртами, которые различаются числом двойных связей в стероидном ядре и строением боковой цепи. Ниже показано строение некоторых из этих липидов: 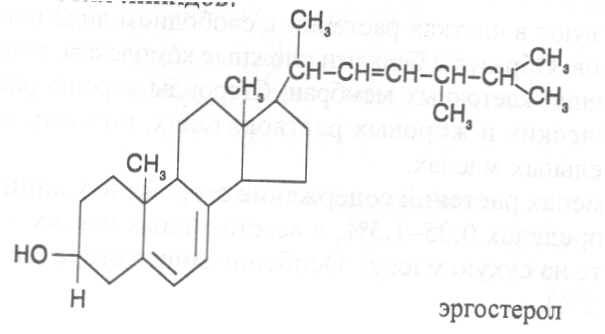 Стероидные производные входят в состав многих биологически активных соединений, играющих важную роль в жизнедеятельности как растений, так и животных организмов: гормонов, сердечных гликозидов, гликоалкалоидов, желчных кислот, витаминов. 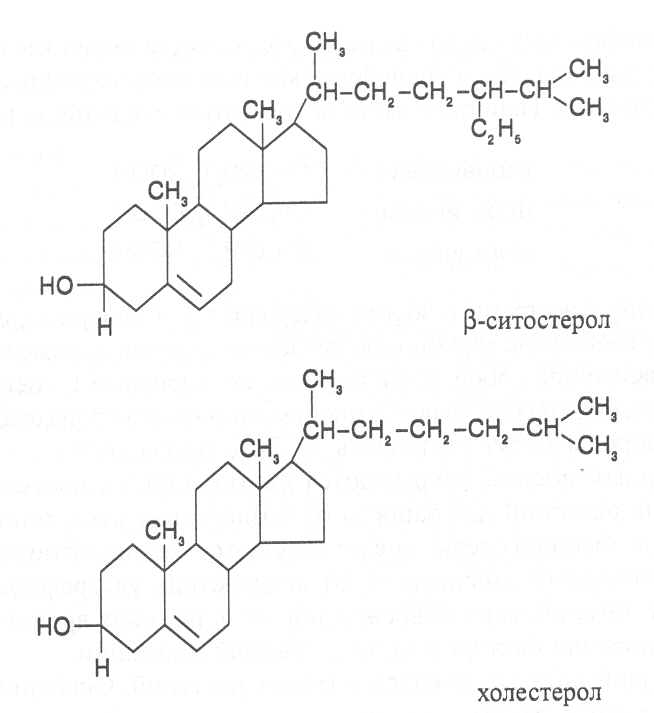 3.5. Строение, свойства и биологические функции восков. К воскам относятся сложные эфиры высокомолекулярных одноатомных спиртов и карбоновых кислот, имеющие твёрдую или жидкую кон-систенцию. В состав таких эфиров очень часто входят спирты: цетиловый СН3(СН2)₁₄СН₂ОН цериловый СН3(СН2)₂₄СН₂ОН мирициловый СН3(СН2)₂₉СН₂ОН Из карбоновых кислот в состав эфиров воска входят как обычные жирные кислоты, так и специфические высокомолекулярные карбоновые кислоты. Наиболее часто встречаются следующие кислоты: карнаубовая СН₃(СН₂)₂₂СООН церотиновая СН₃(СН₂)₂₅СООН монтановая СН₃(СН₂)₂₇СООН Наряду с эфирами в восках содержатся углеводороды, ацилглицерины, свободные карбоновые кислоты и спирты, а также терпены, представляющие собой углеводороды, построенные из нескольких молекул изопрена (см. главу «Вещества вторичного происхождения»). Уг-леводороды могут составлять 20-70% массы воска. Твёрдым воском покрываются листья, стебли, цветки, плоды и семена растений для защиты от воздействия неблагоприятных факторов внешней среды: предохранение от излишней потери воды или чрезмерного смачивания, от воздействия ультрафиолетовых лучей и механических повреждений, от поражения вредителями и проникновения бактериальной и грибной инфекции. Жидкий воск содержится в тканях растений. Особенно много его в оболочках плодов и семян, где он во взаимодействии с твердым воском образует структурную основу покровной ткани. Содержание воска в оболочках семян обычно находится в пределах 0,01-0,2%, в оболочках плодов - 3-18% (в расчёте на сухую массу). При повреждении воскового налёта на поверхности плодов и овощей во время транспортировки и хранения они быстрее поражаются инфекцией и подвергаются порче. Известны растения (в основном тропического происхождения), способные накапливать много воска, который используется для про-мышленной переработки (изготовление свечей, мастики, лекарственных средств). Воск может откладываться толстым слоем на поверхности листьев (восконосная пальма карнауба), стволов (пальмы из рода Ceroxylon), плодов (мирика восконосная), а также в тканях стеблей (лангсдорфия подземная). Особой ценностью обладает воск, образующийся в семядолях растений хохобы (симмондсия китайская), который имеет жидкую консис-тенцию и используется в медицинских целях. Вопросы для повторения. 1. Каковы основные группы липидов и какие они выполняют функции в организмах? 2. Из каких жирных кислот и ацилглицеринов образуются молекулы жиров? 3. В чём состоят особенности твёрдых и жидких жиров, а также жиров разных растений? 4. Какие жирные кислоты называют незаменимыми и почему? 5. Для чего и как используются показатели, называемые числами жиров? 6. Какие химические изменения происходят в процессах прогоркания и высыхания жиров? 7. Как классифицируют растительные масла по способности к высыханию? 8. Каковы структурные и функциональные особенности различных групп фосфолипидов и гликолипидов? 9. Какие известны разновидности стероидных липидов? 10. В чём состоят химические и биологические особенности воска у различных видов растений? 11. В каком количестве содержатся различные группы липидов в растительных продуктах? 12. Как влияют липиды на качество растительной продукции? Резюме по модульной единице 3. К липидам относятся жиры, фосфолипиды, гликолипиды, стероидные липиды и воска. Все они преставляют собой гидрофобные вещества, растворимые в неполярных органических растворителях. Важнейшие компоненты липидов – насыщенные и ненасыщенные жирные кислоты. Полиненасыщенные жирные кислоты с двумя и более двойными связями не могут синтезироваться в организмах человека и животных, их называют незаменимыми жирными кислотами. Основными источниками этих кислот для указанных организмов служат растительные жиры, которые называют маслами в связи с тем, что в их составе преимущественно содержатся остатки ненасыщенных жирных кислот, имеющих низкие температуры плавления и обусловливающие жидкую консистенцию растительных жиров. Жиры представляют собой смесь сложных эфиров глицерина и жирных кислот, называемых ацилглицеринами. Состав ацилглицеринов растительных масел зависит от вида и органа растений, а также от условий их выращивания. Свойства жиров оценивают с помощью показателей, называемых числами жиров (кислотное число, йодное число, число омыления). Жиры откладываются в растениях как запасные вещества и поэтому влияют на качество растительной продукции. Фосфолипиды включают остатки глицерина, нысыщенных и ненасыщенных жирных кислот, а также азотистых оснований и некоторых соединений. Они участвуют в формировании клеточных мембран. Фосфатидилэтаноламины и фосфатидилхолины входят в различные клеточные мембраны и откладываются в семенах растений в качестве запасных веществ. Фосфатидилсерины и фосфатидилинозиты в большом количестве содержатся в составе хлоропластных и митохондриальных мембран, фосфатидилглицерины – в составе хлоропластных мембран. Фосфолипиды участвуют также в регуляторных процессах в составе клеточных мембран. Гликолипиды состоят из остатков глицерина, остатков линоленовой и других жирных кислот, а также остатков галактозы и сульфоглюкозы. Много их содержится в составе хлоропластных мембран. Стероидные липиды представлены производными циклопентанопергидрофенантрена, содержатся в свободной форме и в виде гликозидов. Они участвуют в построении клеточных мембран, эргостерол является провитамином D. Многие стеролы относятся к физиологически активным веществам. Воски образуются из высокомолекулярных одноатомных спиртов и специфических высокомолекулярных карбоновых кислот. В состав восков также входят углеводороды, ароматические и красящие вещества. Воском покрыты поверхности листьев, цветков, семян, плодов, стволов и ветвей растений, они образуют структуру семенных и плодовых оболочек. Тестовые задания к лекции 1. Тесты № 1-28. Лекция 2. Строение, свойства и биологические функции аминокислот, нуклеотидов, белков и витаминов. Аннотация. В данной лекции даётся биохимическая характеристика протеи-ногенных и других аминокислот, пуриновых и пиримидиновых нуклеотидов, основных групп белков и витаминов. Раскрывается биологическая роль этих веществ в растительных и других организмах. Отмечается влияние белков, аминокислот и витаминов на качество растительной продукции. Даются сведения о содержании белков и витаминов в сельскохозяйственных растениях и получаемой из них растительной продукции. Ключевые слова: протеиногенные аминокислоты, незаменимые аминокислоты, меланоидины, меланины, пуриновые нуклеотиды, пиримидиновые нуклеотиды, нуклеозиды, дифосфат- и трифосфатпроизводные нуклеотидов, полипептидная теория строения белков, первичная, вторичная, третичная и четвертичная структура белков, пептидная связь, гидрофобное ядро белковой молекулы, нативная конформация белковой молекулы, денатурация белков, фибриллярные и глобулярные формы белковых молекул, протеины, протеиды, альбумины, глобулины, проламины, глютелины, гликопротеиды, липопротеиды, нуклеопротеиды, гистоны, полноценные и неполноценные белки, биологическая ценность белков, водорастворимые и жирорастворимые витамины, провитамины, антивитамины. Рассматриваемые вопросы:
2. Строение, свойства и биологические функции нуклеотидов. 3. Строение, свойства и биологические функции белков. 4. Строение, свойства и биологические функции витаминов. Модульная единица 4. Аминокислоты, нуклеотиды и белки. Цели и задачи изучения модульной единицы. Изучить строение, свойства и биологические функции аминокислот, нуклеотидов, витаминов и основных групп белков. Научить студентов использовать сведения об аминокислотах, нуклеотидах, белках и витаминах при оценке качества растительной продукции. 4.1. Строение, свойства и биологические функции аминокислот. Аминокислоты - это первичные азотистые вещества растений, которые синтезируются с использованием минерального азота, поступающего главным образом из почвы. В молекулах аминокислот имеются карбоксильные и аминные группировки, соединённые с органическим радикалом алифатической, ароматической или гетероциклической природы. Если аминокислота содержит одну карбоксильную и одну аминную группу, связанную со вторым углеродным атомом ( α-положение), строение такой аминокислоты можно выразить следующей формулой: 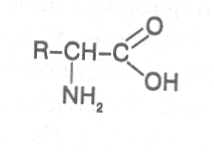 Аминокислоты, имеющие одну карбоксильную и одну аминную группу, принято называть моноамuномонокарбоновымu. У большинства из них аминогруппа находится в α-положении по отношению к атому углерода карбоксильной группы. Однако известны также некоторые аминокислоты, у которых аминогруппа связана с другими углеродными атомами (_b, γ, d и др., см. табл. 1). В организмах также синтезируются аминокислоты с двумя карбоксильными или двумя аминными и другими азотсодержащими группировками. Аминокислоты, содержащие две карбоксильные и одну аминную группы, обычно называют моноамuнодuкaрбоновы.мu, а имеющие две аминные и одну карбоксильную - диаминомонокарбоновымu. Кроме того, аминокислоты различаются по строению радикала R, который может быть представлен неразветвлённой, а иногда и разветвлённой углеродной цепью, ароматическими и гетероциклическими производными. Наряду с аминокислотами важную роль в обмене азотистых веществ играют некоторые иминокислоты (пролин, пипеколииовая кислота и др.), содержащие вторичную аминную группировку (═NH). Они близки по физико-химическим свойствам к истинным аминокислотам и выполняют сходные биологические функции. Важные функции в растительном организме выполняют производные аминокислот – амиды и бетаины, из которых наиболее хорошо изучены аспарагин, глутамин и гликоколбетаин. Аспарагин и глутамин участвуют в построении белковых молекул, являются продуктами обмена многих азотистых веществ. Гликоколбетаин ─ продукт азотного обмена у некоторых растений, служит активным донором метильных групп. Все аминокислоты, за исключением глицина, содержат асимметрические атомы углерода и проявляют оптическую активность. D- и L-формы аминокислот различают по положению водорода и аминогруппы у α-углеродного атома. За эталон сравнения принимаются конфигурации молекул L- и D-серина. Изомеры аминокислот, имеющие расположение в пространстве водорода и аминогруппы у α-углеродного атома такое же, как у L-серина, относят L-ряду, а сходное с конфигурацией молекулы D-серина – к D-ряду. 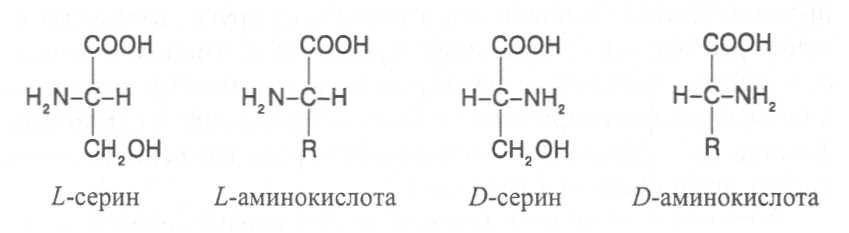 Направление и угол вращения плоскости поляризации света у разных аминокислот и их оптических изомеров зависит от строения радикала R, реакции среды (рН), природы растворителя и растворённых в нём веществ. Подавляющее большинство природных аминокислот синтезируется в организмах в виде L-форм, а D-формы аминокислот встречаются редко, чаще всего в клетках микроорганизмов. При химическом синтезе образуется смесь L- и D-изомеров аминокислот. Ферментные системы растений, человека и животных специфически приспособлены катализировать биохимические реакции, происходящие с участием L-изомеров аминокислот, и не способны к превращениям D-изомеров, которые даже могут ингибировать биохимические процессы в организме. В опытах установлено, что только метионин может усваиваться организмами человека и животных как в L-форме, так и D-форме. Первые аминокислоты были открыты в начале XIX века, а к концу этого века уже были выделены и изучены почти все аминокислоты, входящие в состав белков. В настоящее время известно более 200 аминокислот. Важнейшая биологическая роль аминокислот - построение белковых молекул. Аминокислоты, участвующие в синтезе белков, принято называть протеиногенными, их насчитывается 18. Кроме того, в синтезе белков принимают участие два амида - аспарагин и глутамин. После синтеза белковой молекулы в ней могут происходить модификации радикалов некоторых аминокислот, поэтому при анализе состава белков, кроме протеиногенных, обнаруживают некоторые другие аминокислоты (оксипролин, оксилизин и др.). Аминокислоты, не участвующие в синтезе белков, являются важными метаболитами, с участием которых происходит синтез протеиногенных аминокислот, а также всех других азотистых веществ растительного организма: нуклеотидов, амидов, азотистых оснований, алкалоидов, некоторых липидов, многих витаминов, хлорофилла, фитогормонов (ауксинов, цитокининов), некоторых фитонцидов. Строение и биологическая роль важнейших аминокислот представлены в таблице 1. Растения и природные формы микроорганизмов способны синтезировать все необходимые им аминокислоты из других органических веществ, тогда как организмы человека и животных не способны к синтезу некоторых аминокислот, входящих в состав белков. Эти аминокислоты называют незаменимыми и они должны поступать в организм с пищей. Для взрослого человека незаменимыми являются 8 аминокислот: лизин, триптофан, метионин, треонин, лейцин, валин, изолейцин, фенилаланин. Для детей и некоторых групп животных незаменимыми также являются аргинин, гистидин и цистеин. При недостатке незаменимых аминокислот ослабляется синтез белков, что может быть причиной тяжелых заболеваний. А их недостаток в растительных кормах снижает выход животноводческой продукции в расчете на единицу массы затраченного корма, в результате чего повышается ее себестоимость. В целях составления правильного пищевого рациона для каждого вида организмов с учетом возрастного и физического состояния определены ежедневные нормы потребления незаменимых аминокислот. В среднем для человека они составляют, г: валин–5,0, лейцин–7,0, изолейцин –4,0, лизин–5,5, триптофан–1,0, треонин–4,0, метионин–3,5, фенилаланин –5,0. Чаще всего в кормах сельскохозяйственных животных в недостаточном количестве содержатся такие незаменимые аминокислоты, как лизин, триптофан и метионин. Для балансирования кормов по содержанию этих аминокислот разработаны промышленные способы их получения. В связи с тем, что лизин и триптофан усваиваются животными только в виде L-изомеров, то для производства кормовых препаратов указанных аминокислот применяют микробиологический синтез, при котором реализуется природный механизм образования L-изомеров аминокислот. Поскольку метионин может усваиваться животными в виде D- и L-форм, то для его промышленного получения используется менее затратный химический синтез, дающий рацемическую смесь оптических изомеров этой аминокислоты. Содержание свободных аминокислот в растениях зависит от вида органа или ткани, возраста растений, внешних условий и особенно подвержено большим изменениям в зависимости от интенсивности протекания тех биохимических процессов, которые сопряжены с их потреблением (синтез белков, нуклеиновых кислот и других азотистых веществ). Концентрация аминокислот повышается при ослаблении ростовых процессов, недостатке питательных элементов, избыточном азотном питании, усилении процессов распада белков при старении растений или прорастании семян. Концентрации отдельных аминокислот могут возрастать в результате метаболитных нарушений в организме и под воздействием стрессов. Так, например, при вододефицитном стрессе в клетках растений происходит накопление аминокислоты пролина, а при избыточном аммонийном питании – накопление аспарагина, глутамина и аргинина. 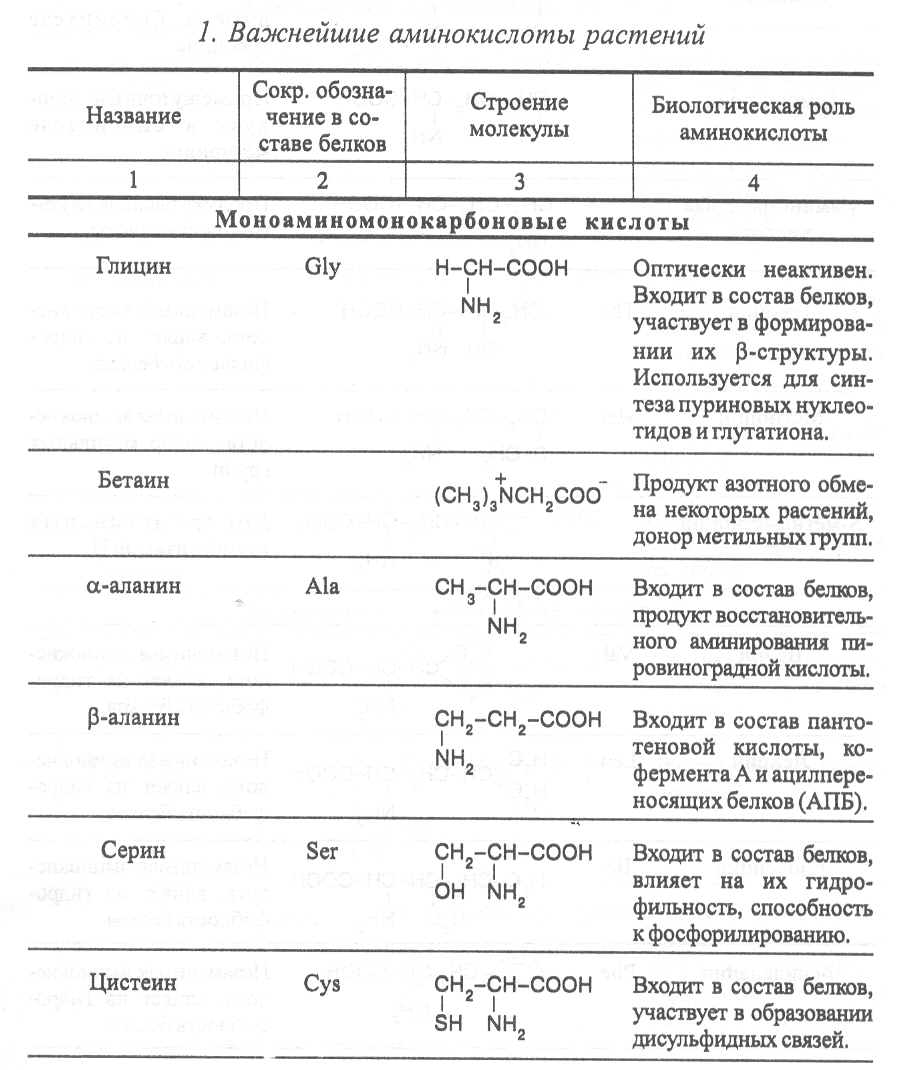 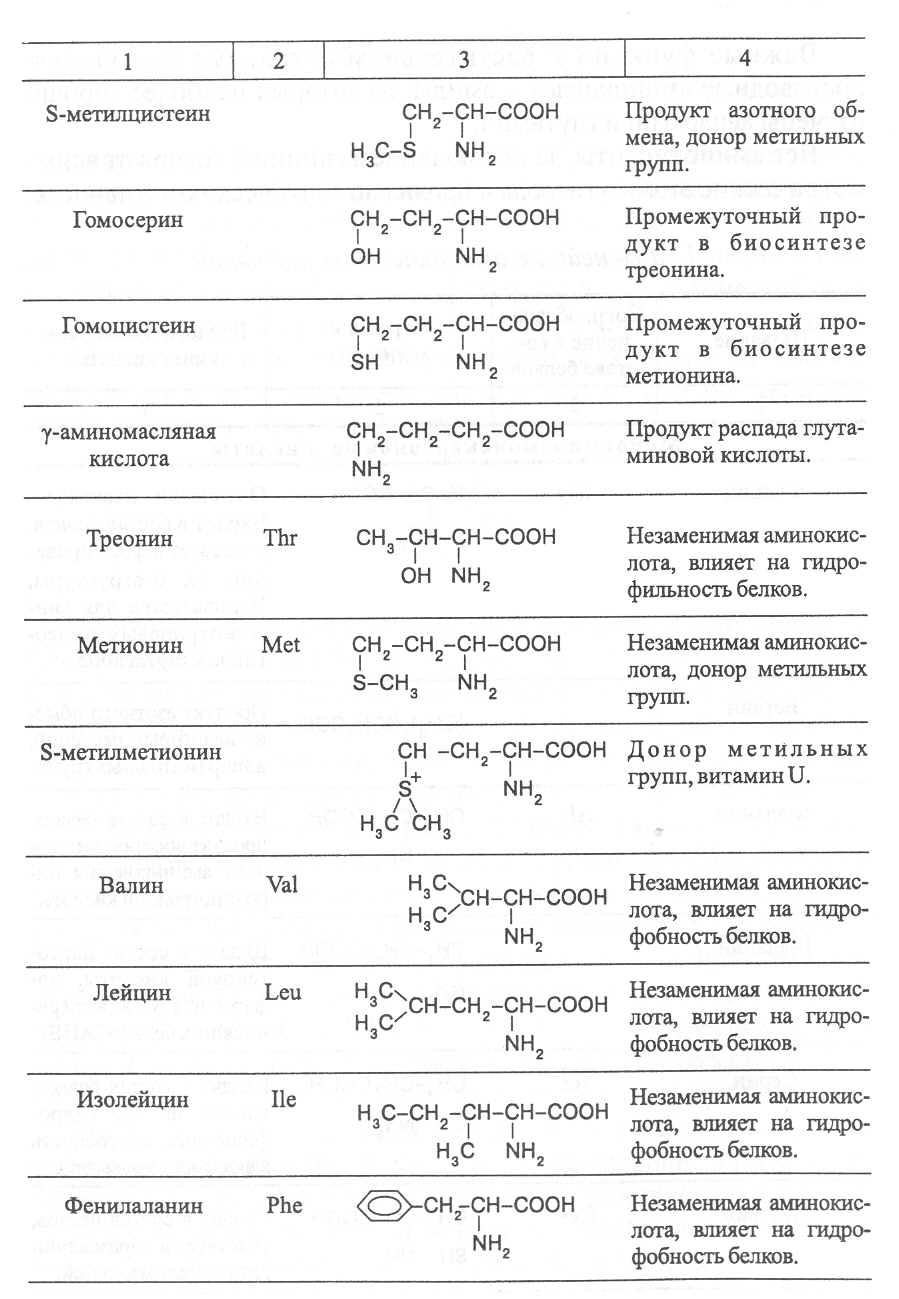  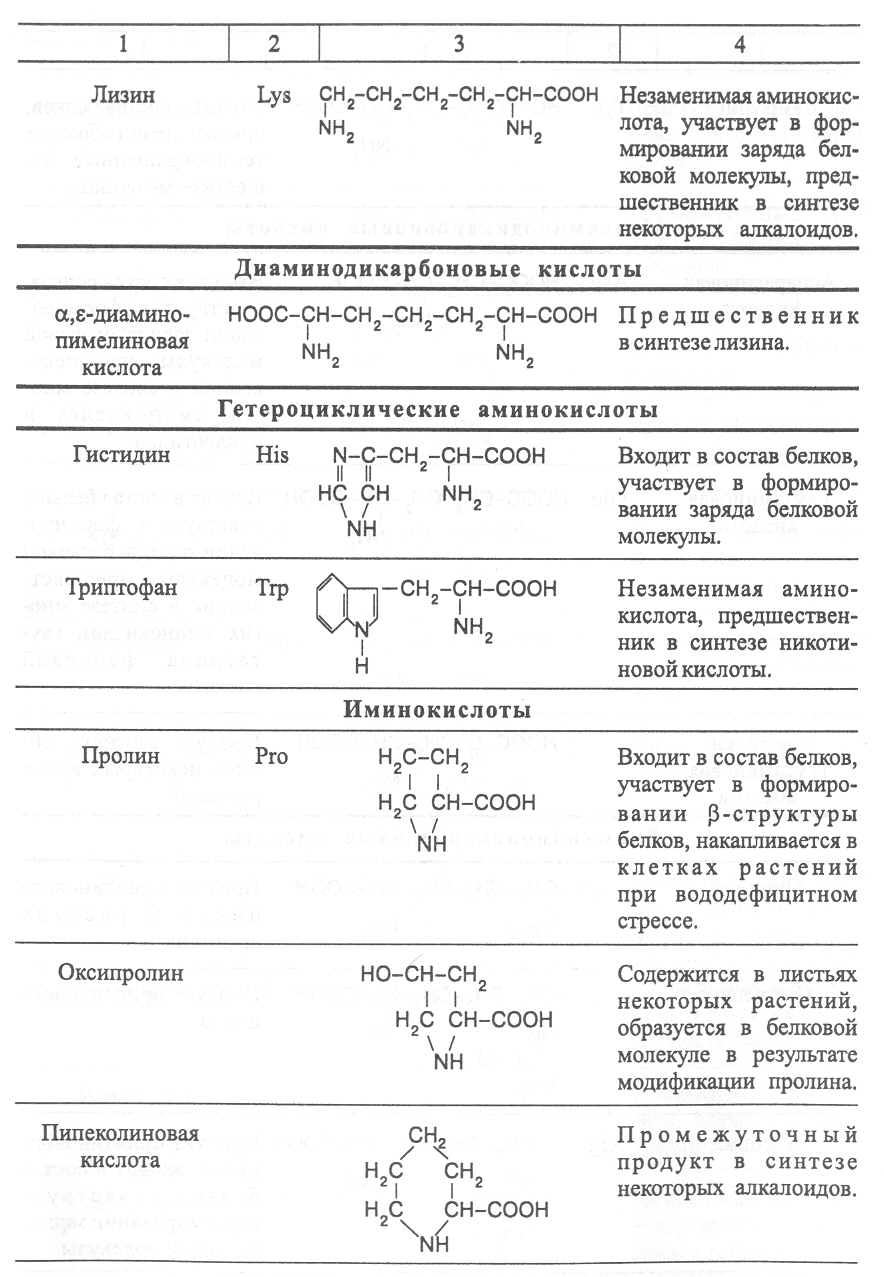 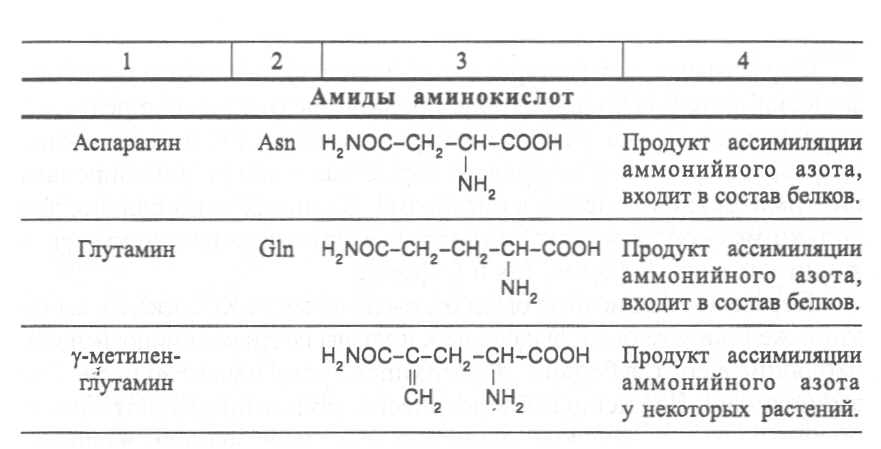 |