Биофизика. Биофизика мембран. Навчальнометодичний посібник для викладачів для проведення занять студентів зі спеціальностей 12010001 Лікувальна справа
 Скачать 6.62 Mb. Скачать 6.62 Mb.
|

|
Завдання: 1.1. Розгляньте рис. 1. Шляхом порівняння з калібрувальним сигналом, виміряйте наступні величини: А) величину потенціалу спокою Б) амплітуду потенціалу дії В) тривалість потенціалу дії. 1.2. Вкажіть, які іонні зрушення обумовлюють фазу деполяризації потенціалу дії. 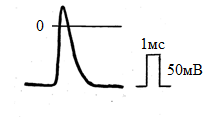 Рис. 1 2. Вплив заміни всередині аксона іонів калію іонами натрію на потенціал дії. Гігантське нервове волокно помістили в ізотонічний сольовий розчин іонів натрію. У відповідь на електричне подразнення в ньому виник потенціал дії (рис.2 а). Після цього ¼ частина іонів калію, що знаходяться всередині волокна замінили іонами натрію і знову записали потенціал дії (ріс.2б). Потім замінили всередині волокна ½ частина іонів калію іонами натрію, записавши потенціал дії. Завдання: 2.1. Розгляньте мал. 2 а - в і проаналізуйте зміна амплітуди і тривалості потенціалу дії. 2.2. Дайте пояснення цьому явищу з точки зору мембранної теорії збудження.  Рис. 2 Варіант 2 1. Вимірювання потенціалу спокою і потенціалу дії одиночного волокна смугастого м'яза. Мікроелектрод ввели всередину м'язового волокна і зареєстрували величину мембранного потенціалу спокою. При подразненні волокна в ньому виник потенціал дії, який також був зареєстрований. Для вимірювання параметрів потенціалу спокою і потенціалу дії записаний калібрувальний сигнал (прямокутний імпульс, амплітуда якого дорівнює 50мВ, а тривалість 1 мс.) Завдання: 1.1. Розгляньте рис.1 та, шляхом порівняння з калібрувальним сигналом, виміряйте наступні величини: А). величину потенціалу спокою; Б). амплітуду потенціалу дії; В). тривалість потенціалу дії. 1.2. Вкажіть іонний механізм фази реполяризації потенціалу дії.  Рис.1 2. Вплив іонів натрію на виникнення потенціалу дії Нерв раку, який містить два нервових волокна, дратували нервовими імпульсами. Виниклі при цьому потенціали дії зареєстрували. Спочатку нерв був занурений в фізіологічний розчин (крива а). Потім його перенесли в розчин, з якого були виключені іони натрію і замінені іонами холіну, не проникаючими через мембрану. Записи «б» - «е» зроблені через рівні проміжки часу після заміни натрію холін. Завдання: 2.1. Розгляньте мал. 2 і проаналізуйте зміна амплітуди потенціалу дії. 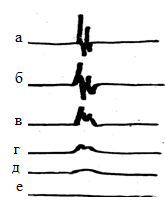 Рис.2 2.2. Поясніть, чому виключення з фізіологічного розчину іонів натрію перешкоджає виникненню потенціалу дії. IV. Контроль знань (письмове опитування з теми) 15 хв. 1. Іонний механізм деполяризації мембрани під час потенціалу дії полягає в дифузії іонів: А. калію зсередини клітини назовні Б. калію зовні клітини всередину В. натрію і калію зсередини клітини Г. натрію зсередини клітини назовні Д. натрію зовні всередину клітини 2. Потенціал електрохімічного рівноваги іонів натрію розраховується за умови, що мембрана: А. непроникна для іонів при порушенні Б. непроникна для пасивного перенесення іонів В. проникна тільки для іонів натрію Г. непроникна для іонів натрію Д. переносить насосами іони натрію 3. Фаза реполяризації мембрани при потенціалі дії пов'язана з: А. збільшенням проникності мембрани для іонів натрію Б. зниженням проникності мембрани для іонів калію В. збільшенням проникності мембрани для іонів калію Г. інактивацією калієвих і натрієвих каналів мембрани Д. зникненням градієнта концентрації іонів натрію 4. Дифузія іонів натрію через мембрану під час потенціалу дії припиняється внаслідок: А. насичення клітини іонами натрію Б. інактивації натрієвих каналів В. зникнення градієнта концентрації іона Г. зміни напрямку градієнта іона Д. активації натрій-калієвого насоса 5. При деполяризації мембрани під час потенціалу дії проникність мембрани для іонів натрію: А. поступово знижується до нуля Б. різко знижується В. різко збільшується Г. трохи збільшується Д. трохи знижується 6. Однією з причин фази реполяризації потенціалу дії є: А. збільшення проникності мембрани для іонів калію Б. збільшення проникності мембрани для іонів натрію В. збільшення проникності мембрани для натрію і калію Г. зниження проникності мембрани для іонів калію Д. зниження проникності мембрани для іонів натрію та калію 7.Причиною активації натрієвих каналів служить: А. деформація В. реполяризация Д. реверсія Б. рефрактерність Г. деполяризація 8. Вкажіть дві причини реполяризації мембрани під час потенціалу дії: А. інактивація переносу іонів натрію і калію через мембрану Б. активація натрієвих і калієвих каналів мембрани нервового волокна В. інактивація натрієвих каналів і збільшення проникності для калію Г. інактивація калієвих каналів і збільшення проникності для натрію Д. активація натрієвих каналів і зниження проникності мембрани для калію 9. Вкажіть правильну послідовність фаз потенціалу дії: А. деполяризація-ремісія-реверсія Б. реполяризация-реверсія-деполяризація В. деполяризація-реполяризация-реверсія Г. деполяризація-реверсія-реполяризация Д. реполяризация-деполяризація-ревміссія 10. Проведення імпульсів по нервовому волокну може здійснюватися: А. в обидва боки від місця виникнення, але не в зворотному напрямку Б. в обидва боки від місця виникнення і в зворотному напрямку Г. в одну сторону від місця виникнення, але можливо у зворотному напрямку Д. ізольовано і тільки в одну сторону від місця виникнення 11. Швидкість проведення нервового імпульсу по міелінізірованние нервовому волокну: А. не залежить від його діаметра: вона завжди постійна Б. залежить від діаметра: чим тонше волокно, тим швидкість більше В. не залежить від діаметра: вона визначається тільки довжиною волокна Г. залежить від діаметра: чим товще волокно, тим швидкість більше Д. не залежить від діаметра: вона визначається амплітудою імпульсу 12. Проведення нервових імпульсів в неміелінізірованних нервових волокнах здійснюється за рахунок: А. місцевих електричних струмів між ділянками волокна Б. стрибків потенціалу між перехопленнями Ранвьє В. стрибка потенціалу від початку в кінець волокна Г. зміни мембранного потенціалу одночасно у всьому волокні Д. включення активного транспорту іонів у проведення 13. Сальтоторное проведення нервового імпульсу: А. проведення його від точки до точки в міелінізірованние волокна Б. проведення його від точки до точки в неміелінізірованние волокні В. проведення його стрибками в міелінізірованних волокнах Г. проведення його стрибками в неміелінізірованних волокнах Д. стрибок імпульсу від початку до кінця нервового волокна 14. Найменша швидкість проведення нервового імпульсу характерна для: А. товстих неміелінізірованних волокон Б. тонких неміелінізірованних волокон В. товстих міелінізірованних волокон Г. тонких міелінізірованних волокон Д. будь-яких волокон, покритих мієліном 15. Найбільша швидкість проведення нервового імпульсу характерна для: А. тонких міелінізірованних волокон Б. тонких неміелінізірованних волокон В. будь-яких волокон, не покритих мієліном Г. товстих неміелінізірованних волокон Д. товстих міелінізірованних волокон 16. Вкажіть причину, по якій зворотне проведення нервового імпульсу по нервовому волокну неможливо: А. активація натрій-калієвого насоса Б. інактивація натрій-калієвого насоса В. інактивація калієвих каналів Г. інактивація натрієвих каналів Д. активація калієвих каналів мембрани 17. Вкажіть, чим калієві канали нервового волокна відрізняються від натрієвих каналів: А. відсутністю іонної специфічності Б. відсутністю інактіваціонних воріт В. відсутністю активаційних воріт Г. зарядом гідрофільних груп білка в каналі Д. способом управління воротами каналу 18. Під час фази деполяризації потенціалу дії більшість натрієвих каналів: А. спочатку відкриваються, а потім закриваються Б. спочатку закриваються, потім інактивуються В. спочатку відкриваються, потім інактивуються Г. спочатку інактивуються, потім відкриваються Д. перебувають у відкритому стані завжди 19. У стані спокою більшість натрієвих каналів мембрани: А. відкриті В. активовані Д. деполярізовані Б. закриті Г. інактивовані 20. Такий фази потенціалу дії не існує: А. реверсії потенціалу В. деполяризації Б. реполяризації Г. ремісії Д. слідової деполяризації V. Підведення підсумків заняття, домашнє завдання 10 хв.: Тема наступного заняття: «Біофізика м’язового скорочення» Питання для підготовки до заняття: 1. Структура скелетного м’яза і м’язового волокна. Актинові та міозинові міофібрили. Поняття про саркомер. 2. Роль потенціалу дії у виникненні м’язового скорочення. 3. Молекулярний механізм м’язового скорочення. 4. Взаємодія міозину, актину і кальцію в процесі м’язового скорочення. 5. Енергетика м’язового скорочення. 6. Зв’язок швидкості скорочення м’яза з прикладеним до м’яза навантаженням. Рівняння Хіла. 7. Види скорочення скелетного м’язу. 8. Взаємодія м’язів з кістковою системою. ЛІТЕРАТУРА: 1. Основи біофізики / / Навчальний посібник для студентів медичного факультету - Запоріжжя, ЗДМУ, 2011. 2. Ємчик Л.Ф., Кміт Я.М. Медична і біологічна фізика- Львів: Світ, 2003.-с.171-180. ТЕМА ЗАНЯТТЯ 6: БІОФІЗИКА М'ЯЗОВОГО СКОРОЧЕННЯ АКТУАЛЬНІСТЬ ТЕМИ: Рух – одне з основних і загальних властивостей всього живого. Біофізичні механізми, які полягають в основі руху, дуже різноманітні. Для студентів медичного факультету найбільш актуальними є м'язові скорочення, як приклад високоспеціалізованого руху. Різні типи м'язів своїми скороченнями створюють і формують рух кінцівок, всього тіла та його частин (скелетні м'язи), внутрішніх органів і судин (гладенькі м'язи), серця (серцевий м'яз). Одну з найважливійших функцій у життєдіяльності організма відіграють скелетні м'язи. За нашего часу їх діяльність встановлена і на молекулярном, і на клітинном рівні. У подальшому навчанні (насамперед, у ході нормальної фізіології) вивчення біофізики м’язового скорочення слугує основою для розуміння фізіології м'язів. ЦІЛІ ЗАНЯТТЯ: Характеризувати структуру м'яза та м'язового волокна; Обгрунтувати роль потенціалу дії у м'язовому скороченні; Трактувати молекулярний механізм м'язового скорочення; Встановити зв'язок між швидкостью скорочення м’яза і прикладеним до нього навантаження; Відокремити різні види м’язового скорочення; Характеризувати взаємодію м'язів з кістковою системою. МАТЕРІАЛИ МЕТОДИЧНОГО ЗАБЕЗПЕЧЕННЯ: 1. Матеріали для основної частини заняття: таблиці, схеми, малюнки, проблемні теоретичні питання зі збірника «Робочий зошит для самостійної роботи на практичних заняттях для студентів медичного факультету» 2. Матеріали для контролю знань:тестові питання для стандартизованого контролю знань студентів. 3. Матеріали для самопідготовки студентів:рекомендовані навчальні посібники, методичні вказівки для самостійної роботи студентів. ТЕОРЕТИЧНІ ВІДОМОСТІ: Скелетне м'язове волокно являє собою многоядерную клітину якої довжина дорівнює довжині м'язи в цілому. М'язові волокна оточені поляризованою плазматичної мембраною. Мембранний потенціал спокою скелетного м'язово-го волокна становить близько - 90мВ. Кожне м'язове волокно включає в себе міофібрили, які безпосередньо беруть участь в м'язовому скороченні. Міофібрили занурені в саркоплазму, в якій знаходяться звичайні внутрішньоклітинні органели. Основними іонами рідини саркоплазми є катіони калію, аніони фосфату. Відмінною рисою саркоплазми м'язового волокна є наявність в ній безлічі мітохондрій, що синтезують велику кількість АТФ. У саркоплазме є також дві спеціалізовані мембранні системи: Т-система і саркоплазматический ретикулум. Т-система являє собою впячивания поверхневої мембрани в поперечному напрямку всередину м'язового волокна. Вона контактує з саркоплазматическим ретикулумом - системою, всередині якої містяться у високій концентрації іони кальцію. Міофібрили складаються з менших субодиниць - миофиламентов, які представляють собою білкові молекули (близько 1500 миозинових і 3000 актінових филаментов). Міозіновие філаменти є більш товстими (15 нм), в порівнянні з актиновими (10 нм). Міозинові і актинові філаменти частково перекриваються, в результаті чого міофібрили в мікроскопі виглядають як сукупність світлих і темних дисків. Світлі диски містять тільки актинові філаменти і називаються ізотропним (I-диски). Темні диски анізотропнимі (А-диски), представлені міозіновимі филаментами і розташованими між ними кінцями актинових філаментів. Від поверхні миозинових филаментов уздовж всієї їх довжини (за винятком центральної частини) відходять поперечні містки. У м'яза, який на скорочується кінці миозинових і актинових філаментів зазвичай лише слабо перекриваються на кордоні між А- і I-дисками. Ця зона перекривання виглядає в світловому мікроскопі тимніше центральної Н - зони, в якій немає актинових ниток. В середині саркомера (в центрі Н-зони) видна тонка темна М-лінія - мережа опорних білків, які утримують міозіновие філаменти в складі єдиного пучка. Кінці актинових волокон прикріплюються до, так званої, Z-мембрани, від якої вони тягнуться в обох напрямках до місць перекриття з міозіновимі филаментами. Z - мембрана знаходиться поперек кожної міофібрили і з'єднує все міофібрили м'язового волокна один з одним. Отже, м'язове волокно в цілому має такі ж світлі і темні диски, як і окремі міофібрили. Ці диски утворюють повторювані структурні елементи - саркомери, які надають скелетної м'язі смугастість. Саркомер - частина міофібрил (або м'язового волокна в цілому), яка лежить між двома сусідніми Z-мембранами. Управління скорочувальної активністю скелетних м'язів здійснюється аксонами рухових нервових клітин – мотонейронів, кожен з яких іннервує групу м'язових волокон (нейромоторная одиниця). Хакслі і Хансон показали, що під час м'язового скорочення молекули актину і міозину не змінюють своєї довжини. Вони лише переміщуються один вздовж одного, в результаті чого довжина окремих міофібрил і м'яза в цілому зменшується (теорія "ковзающих ниток»). Скороченню м'язи передує ряд подій, що відбуваються в її м'язових волокнах і запускають процес скорочення. 1. М'язове волокно активується імпульсами, що приходять по нервовому волокну; 2. В плазматичній мембрані м'язового волокна виникає потенціал дії; 3. Потенціал дії деполяризує мембрану м'яза і потенціал дії переміщується вздовж мембрани; 4. Деполяризация мембрани переміщується вглиб м'язового волокна по канальцям Т-системи і саркоплазматичного ретикулума. Це викликає вивільнення з саркоплазматичного ретикулума через потенціал-залежні кальцієві канали великої кількості іонів кальцію в саркоплазму; 5. Іони кальцію ініціюють взаємодію між актиновими і міозіновимі филаментами, змушуючи їх ковзати один щодо одного, що і викликає процес скорочення м'яза; 6. Через короткий час іони кальцію відкачуються з саркоплазми в саркоплазматический ретикулум шляхом активного транспорту (роботи кальцієвого насоса). Видалення іонів кальцію з саркоплазми призводить до припинення скорочення. У стані спокою актинові філаменти кожного саркомера, прикріплені кінцями до Z-пластинок, трохи перекривають міозинові філаменти. При скороченні м'язового волокна актинові філаменти підтягуються в проміжки між міозіновимі филаментами так, що починають перекривати їх практично на всьому протязі саркомера. Z-мембрани підтягують актинові філаменти до кінців міозивих филаментов, скорочення м'яза здійснюється за механізмом "ковзающих ниток». В результаті довжина кожного саркомера і м'яза в цілому зменшується. Молекулярна структура миозинових і актинові філаментів в даний час детально вивчена. Міозінових филамент складається з молекул міозину (білок з М=500000). Кожна з цих молекул сформована шістьма поліпептидними ланцюгами: двома важкими і чотирма легкими. Два важкі ланцюги згорнуті навколо один одного, формуючи подвійну спіраль. Один кінець кожної з важких ланцюгів згорнуть в грушеподібну глобулярную структуру, яка називається головкою міозину. Їх складовими частинами є також чотири легкі ланцюги міозину. Головки міозину здатні в присутність актину каталізувати реакцію гідролізу АТФ. Подовжена частина спіралі називається хвостом. Частина спіралі кожної молекули міозину разом з головкою формує попе-річковий місток. Двісті або більше молекул міозину, пов'язані разом, формують структуру миозинового філамента, причому хвости миозинових молекул спрямовані до середини саркомера, а головки орієнтовані так, що можуть сприяти руху. Більш тонкі актинові філаменти також мають складну структуру. Вони сформовані з трьох білкових компонентів: актину (білок з М=42000) і двох кальцій-чутливих регуляторних білків: тропоміозіна і тропоніна. У кожному актиновому филаменті дві молекули актину згорнуті, формуючи спіраль, на поверхні якої розташовані активні центри - ділянки, до яких можуть прикріплятися поперечні містки молекул міозину при скороченні м'яза. Молекули тропоміозина розташовані в жолобках, утворених спірально закрученими молекулами актину і в стані спокою прикривають активні центри актинових молекул, запобігаючи взаємодії між ними і поперечними містками міозину. Молекули тропоніна прилягають до поверхні молекул тропоміозина і мають велику спорідненість до іонів кальцію. При взаємодій- тропоніна з іонами кальцію його молекула змінює свою конформацію так, що як би заштовхує тропоміозин глибше в жолобок між двома актиновими молекулами. При цьому відкриваються активні центри актинових филаментов, і відбувається прикріплення до них поперічкових містків міозину, що призводить до скорочення м'яза. При ініціації м'язового скорочення з саркоплазматичного ретикулума виходять у великій кількості іони кальцію. Вони зв'язуються з молекулами тропоніна, це призводить до конформаційних перебудов його структури, що зумовлює взаємодію між тропонін-тропоміозіном і молекулами актину. В результаті активні центри актиновой спіралі стають відкритими і здатними взаємодіяти з поперечними містками міозину. Головки поперечних містків міозинових молекул приєднуються до активних центрів актинових филаментов. Основну роль у взаємодії поперечних містків міозину з актином грають електростатичні сили і гідрофобні взаємодії. Спочатку головка кожного поперечного містка «підключається» до активного центру актинового філамента під прямим кутом, і зразу ж нахиляється приблизно до кута 450. В результаті розвивається пружний натяг, що зміщує актиновий филамент приблизно на 10 нм. Після цього головка від'єднується від активного центру актинового філамента, і, повертаючись в свою нормальну позицію, формує зв'язок з новим активним центром молекули актину. У цьому процесі затрачується енергія, яка вивільняється в результаті гідролізу молекули АТФ. Такий процес настає знову і знову, доки актинові філаменти не втягнуться практично повністю між міозиновими філаментами, підтягуючи Z-мембрани до кінців міозінових філаментів. Енергетика м'язового скорочення При скороченні м'яза виконується робота, і на це потрібна енергія, джерелом якої служить АТФ. Головки поперечних мостків міозину в присутність молекул актину функціонують як фермент аденозинтрифосфатаза (ATФаза). Ця властивість дозволяє голівці гідролізувати ATФ і використовувати енергію для скорочення. Гідроліз АТФ відбувається в той момент, коли головка поперекового містка приєднана до актиновой молекулі під кутом 450 від'єднується від активного центру і відбувається зміна її конформації. В результаті головка приєднується під кутом 900 до наступного активного центру молекули актину. Таким чином, для кожного "кроку" поперечного містка необхідна одна молекула АТФ. Зв'язок швидкості скорочення м'яза з доданим навантаженням вивчав Хілл, який встановив зворотну пропорційну залежність між навантаженням м'яза і швидкістю її скорочення. Максимальна ефективність скорочення може бути реалізована в тому випадку, якщо м'яз скорочується з помірною швидкістю. Зазвичай максимальна ефективність проявляє тоді, коли швидкість скорочення становить близько 30% максимальної. |