Нормальная физиология. нф теория. Общая физиология возбудимых тканей. Рецептор, нерв, синапс, мышцы
 Скачать 0.68 Mb. Скачать 0.68 Mb.
|

|
Ядерную сумку в виде спирали окружают нервные волокна чувствительного нейрона-первичные рецепторные окончания. В области миотрубок нервные окончания афферентных нейронов гроздевидно ветвятся, образуявторичные рецепторные окончания. Возбуждение от рецепторных окончаний по афферентным волокнам поступает в спинной мозг к мотонейронам, расположенным в передних рогах. Мотонейроны спинного мозга принято подразделять на альфа- и гамма-мотонейроны (так как их аксоны относятся к А-альфа и А-гамма нервным волокнам). Возбуждение от альфа-мотонейронов поступает к экстрафузальным мышечным волокнам, вызывая их сокращение - тонус восстанавливается. Избыточное сокращение экстрафузальных мышечных волокон приводит к растяжению сухожильных рецепторов Гольджи, так как они прикрепляются к мышце последовательно. В них возникает возбуждение, которое поступает к тормозным вставочным нейронам спинного мозга, а от них к альфа-мотонейронам. Активность альфа-мотонейронов при этом снижается, уменьшается импульсация, идущая от них к экстрафузальным мышечным волокнам, тонус несколько снижается. Рассмотренные выше механизмы поддержания мышечного тонуса осуществляются на уровне спинного мозга, поэтому такой тонус называется спинальным или простейшим. Спинальный тонус характеризуется очень слабой выраженностью тонического напряжения. Такой тонус не может обеспечить поддержание позы животного и акт ходьбы, но он достаточен для осуществления простейших спинальных рефлексов. Контрактильный тонус имеет рефлекторную природу. Это доказывается тем, что при перерезке передних или задних корешков спинного мозга, иннервирующих конечность, ригидность мускулатуры этой конечности исчезает. Важную роль в возникновении контрактильного тонуса играет дорсальное вестибулярное ядро продолговатого мозга (ядро Дейтерса), которое возбуждается импульсами от рецепторов вестибулярного аппарата. При раздражении вестибулярного ядра ригидность усиливается, а при его разрушении ригидность уменьшается. Полагают, что это ядро оказывает влияние на ретикулярную формацию продолговатого мозга, которая оказывает неодинаковое влияние на нейроны спинного мозга. Раздражение медиальных отделов ретикулярной формации приводит к торможению рефлексов спинного мозга (тормозящая ретикуло-спинальная система), а раздражение латеральных отделов вызывает активацию. 33.Статические и статокинетические рефлексы. Участие продолговатого и среднего мозга в регуляции мышечного тонуса. На уровне ствола мозга осуществляется тонический рефлекс, который направлен на сохранение равновесия тела в пространстве, в статическом состоянии и при движении. 2 группы: 1)статические-поддерживают равновесие при нормальном положении без перемещения тела 2)статокинетические-равновесие при перемещении тела Статические: 1.рефлексы положения-позно-тонические: раздражают лабиринты вестибулярного аппарата, положение головы по отношению к туловищу. Обычно голова устанавливается в положении темени вверх, когда положение изменяется возбуждаются вестибулярные рецепторы, которые приводят к изменению тонуса скелетных мышц: А) голова вверх: усиливается тонус разгибателей передних конечностей и сгибателей задних Б)голова вниз: усиливается тонус разгибателей задних конечностей и разгибателей передних В)повороты головы в сторону: повышается тонус мышц разгибателей обеих конечностей на той стороне куда повернута голова, повышается тонус сгибателей-в противоположную сторону 2.выпрямтельные рефлексы: раздражается лабиринт вестибулярного аппарата; возбуждение от рецепторов шейных мышц; от кожных рецепторов той поверхности тела на которой лежит животное; от сетчатки глаз Фазы выпрямительного рефлекса: 1)поднятие и поворот головы темени вверх 2)выпрямление туловища за счет перераспределение тонуса мышц сгибателей и разгибателей вызывают вставание животного Статокинетические рефлексы: 1.вращательный рефлекс: возникает при вращении тела вокруг оси, поворот глаз в противоположную сторону и быстрое возвращение в исходное состояние 2.лифтные рефлексы: при быстром вертикальном перемещении вверх или вниз: при подъеме вверх-повышается тонус сгибателей, при спуске вниз-повышение разгибателей 3.рефлекс готовности к прыжку-при резком опускании животного вниз: выпрямление передних конечностей и сгибание задних 34.Ретикулярная формация ствола мозга и ее нисходящее влияние на рефлекторную деятельность спинного мозга. Восходящее активирующее влияние ретикулярной формации на кору больших полушарий.???? Ретикулярная формация - совокупность различных нейронов, расположенных на протяжении ствола мозга, оказывающих активирующее или тормозящее влияние на различные структуры центральной нервной системы, тем самым контролируя их рефлекторную деятельность. Ретикулярная формация ствола мозга оказывает активирующее влияние на клетки коры головного мозга и тормозное действие на мотонейроны спинного мозга. Посылая в спинной мозг к его двигательным нейронам тормозящие и возбуждающие импульсы ретикулярная формация участвует в регуляции тонуса скелетных мышц. Ретикулярная формация поддерживает тонус вегетативных центров, интегрирует симпатические и парасимпатические влияния, передает модулирующее влияние от гипоталамуса и мозжечка к внутренним органам. Функции ретикулярной формации Соматодвигательный контроль (активация скелетной мускулатуры), может быть прямым через tr. reticulospinalis и непрямым через мозжечок, оливы, бугорки четверохолмия, красное ядро, черное вещество, полосатое тело, ядра таламуса и даже соматомоторные зоны коры. Соматочувствительный контроль, т.е. снижение уровней соматосенсорной информации — «медленная боль», модификация восприятия различных видов сенсорной чувствительности (слуха, зрения, вестибуляции, обоняния). Висцеромоторный контроль состояния сердечно-сосудистой, дыхательной систем, активности гладкой мускулатуры различных внутренних органов. Нейроэндокринная трансдукция через влияние на нейромедиаторы, центры гипоталамуса и далее гипофиз. Биоритмы через связи с гипоталамусом и шишковидной железой. Различные функциональные состояния организма (сон, пробуждение, состояние сознания, поведение) осуществляются посредством многочисленных связей ядер ретикулярной формации со всеми частями ЦНС. Координация работы разных центров ствола мозга, обеспечивающих сложные висцеральные рефлекторные ответы (чихание, кашель, рвота, зевота, жевание, сосание, глотание и др.). Восходящие и нисходящие влияния. Ретикулярной формации (РФ). Анатомии ЦНС и нейрофизиология В продолговатом мозге и других стволовых отделах мозга (Варолиевом мосту и среднем мозге) расположена особая клеточная структура – ретикулярная формация. В функциональном отношении ретикулярная формация ствола представляет собой единую структуру. Название этой структуры отражает ранние представления о том, что отдельные нейроны ретикулярной формации имеют обширные связи друг с другом и образуют нечто подобное нейропилю, в котором возбуждение распространяется диффузно, подобно тому, как это происходит в нервной системе кишечнополостных. Однако в дальнейшем эти представления не подтвердились. Четких границ между отдельными ретикулярными и не ретикулярными группами клеток не установлено, однако, выделено до 98 ядерных групп, относящихся к ретикулярной формации. Основными ядрами являются: ядра шва и гигантоклеточное ядро продолговатого мозга, центральное и ретикулярное ядра моста. Клетки ретикулярной формации очень разнообразны по форме и величине. Для них характерно наличие значительно разветвленного дендритного дерева и длинных аксонов. Афферентные входы ретикулярная формация получает как по коллатералям восходящих (сенсорных) путей, так и от вышележащих структур, в том числе от коры больших полушарий и мозжечка. Таким образом, ретикулярная формация интегрирует влияние большого числа мозговых структур. В свою очередь сама она оказывает влияние, как на вышележащие, так и на нижележащие структуры. Нисходящие и восходящие волокна ретикулярной формации покидают ядра на всем ее протяжении, не имея четких пространственных разграничений. Кроме того, существуют аксоны, которые Т-образно ветвятся. Одна из ветвей идет вниз, а другая – в верхние отделы головного мозга. Нисходящие волокна образуют ретикулоспинальный тракт. Через ретикулоспинальный тракт ретикулярная формация оказывает влияние, как на двигательную деятельность спинного мозга (осуществление спинальных рефлексов), так и на вегетативную регуляцию (сосудодвигательная, дыхательная, пищеварительная функции). Ретикулярная формация оказывает влияние на соматические и вегетативные центры в двух противоположных направлениях: торможения и возбуждения. Восходящие влияния ретикулярной формации направлены на регуляцию деятельности коры больших полушарий. Большинство ретикулярных волокон достигает коры через переключение в неспецифических ядрах таламуса. Действие восходящих ретикулярных влияний заключается в широкой активации корковых структур. Торможение активности ретикулярной формации ведет к наступлению сна, активация ее приводит к реакции пробуждения. Характерной особенностью нейронов ретикулярной формации является их высокая чувствительность к химическим факторам. Такие факторы как уровень углекислого газа и кислорода, содержание адреналина, ацетилхолина и серотонина, относительно небольшие концентрации фармакологических веществ изменяют активность нейронов ретикулярной формации, а вместе с этим и ее влияние на кору больших полушарий, соматические и вегетативные рефлексы. Неспецифические нисходящие влияния ретикулярной формации Влияния на двигательную деятельность спинного мозга возникают, в основном, при раздражении ретикулярной формации каудальных отделов ствола. Участки, которые создают эти эффекты, в настоящее время довольно хорошо определены, они совпадают с гигантоклеточным ядром ретикулярной формации продолговатого мозга и ретикулярным ядром моста. Эти ядра содержат большие клетки, аксоны которых образуют ретикулоспинальные пути. Первые работы X.Мегоуна показали, что раздражение гигантоклеточного ядра вызывает ослабление в одинаковой мере всех спинальных двигательных рефлексов: и сгибательных, и разгибательных. Поэтому он сделал вывод о том, что нисходящая система, берущая начало в вентрокаудальной части ретикулярной формации, имеет неспецифическую тормозящую функцию. Несколько позже было обнаружено, что раздражение более дорсальных и ростральных ее участков вызывало, наоборот, диффузное облегчающее действие на спинальную рефлекторную деятельность. Микроэлектродные исследования эффектов, возникающих в нейронах спинного мозга при раздражении ретикулярной формации, действительно показали, что ретикулоспинальные влияния могут изменять передачу импульсации почти во всех рефлекторных дугах спинного мозга. Эти изменения оказываются очень глубокими и длительными, даже в том случае, когда ретикулярная формация раздражается всего несколькими стимулами, эффект в спинном мозге сохраняется сотни миллисекунд. Одновременная активация большого количества ретикулоспинальных нейронов, которая имеет место в эксперименте с прямым раздражением ретикулярной формации и приводит к генерализованному изменению рефлекторной деятельности спинного мозга ситуация, конечно, искусственная. В естественных условиях такого глубокого сдвига этой деятельности, вероятно, не происходит; тем не менее, диффузное изменение рефлекторной возбудимости спинного мозга, несомненно, может иметь место при определенных состояниях мозга. Возможность диффузного ослабления рефлекторной возбудимости можно представить себе, например, при сне; такое воздействие приведет к снижению деятельности двигательной системы, характерному для сна. Важно учесть, что ретикулярное торможение захватывает также спинальные нейроны, участвующие в передаче афферентной импульсации в восходящем направлении, следовательно, оно должно ослаблять передачу сенсорной информации в высшие мозговые центры. Синаптические механизмы диффузных влияний ретикулярной формации на нейроны спинного мозга изучены пока недостаточно. Как уже указывалось, эти влияния чрезвычайно длительны; кроме того, ретикулярное торможение устойчиво к действию стрихнина. Стрихнин является специфическим ядом, устраняющим постсинаптическое торможение мотонейронов, вызванное импульсацией от проприорецепторов (рефлекс сухожильного органа) и связанное с выделением медиатора глицина. Нечувствительность диффузного ретикулярного торможения к стрихнину говорит, по-видимому, о том, что ретикулярные тормозящие эффекты создаются действием на спинальные клетки другого медиатора. Гистохимические исследования показали, что некоторое количество волокон в нисходящих путях, идущих из ретикулярной формации, является по своей природе адренэргическим. Однако пока неизвестно, имеют ли эти волокна отношение к диффузным ретикулоспинальным тормозящим эффектам. Если сравнить нисходящие влияния ретикулярной формации на нейронные структуры, регулирующие соматические и висцеральные функции, то можно обнаружить в них определенное сходство. Как сосудодвигательная, так и дыхательная функции ретикулярной формации строятся на сочетании деятельности двух реципрокно связанных между собой групп нейронов, оказывающих противоположное действие на спинальные структуры. Ретикулярные влияния на спинальные двигательные центры также состоят из противоположных, тормозящих и облегчающих компонентов. Поэтому создается впечатление, что реципрокный принцип организации нисходящих проекций представляет собой общее свойство ретикулярных структур; конечный эффект, соматический или вегетативный, определяется только тем, куда направлены аксоны соответствующих ретикулярных клеток. Такое сходство можно отметить и в других особенностях функционирования ретикулярных нейронов. Ретикулярные структуры, регулирующие вегетативные функции, отличаются высокой химической чувствительностью; влияния ретикулярной формации на двигательные центры также легко изменяются под влиянием таких химических факторов, как уровень CO2 в крови и содержание в ней физиологически активных веществ (адреналина). Механизм действия адреналина на ретикулярные нейроны довольно долго вызывал споры. Дело в том, что адреналин даже при его введении непосредственно в мозговую артерию может оказывать на ретикулярные нейроны непрямое действие (путем, например, сужения мозговых сосудов с последующей аноксией мозговой ткани). Однако исследование реакций ретикулярных нейронов в ответ на прямое приложение к ним адреналина через внеклеточный микроэлектрод показало, что некоторые из них действительно являются адреноцептивными. Восходящие влияния Наряду с функциями, которые осуществляются через нисходящие пути, у ретикулярной формации есть не менее существенные функции, которые осуществляются через ее восходящие пути. Они связаны с регуляцией активности высших отделов мозга, главным образом, коры больших полушарий. Данные о том, что ретикулярная формация играет важную роль в поддержании нормальной деятельности коры больших полушарий, были получены еще в тридцатые годы XX века. Бельгийский нейрофизиолог Ф.Бремер (1935), проводя перерезку головного мозга на различных уровнях и наблюдая за функциями отделенных от остальной центральной нервной системы участков головного мозга, обратил внимание на то, что имеется чрезвычайно существенное различие между животным, у которого перерезка проведена по межколликулярному уровню (между передними и задними холмами четверохолмия), и животным, у которого линия разреза прошла между продолговатым и спинным мозгом. Первый препарат был назван Бремером cerveau isolee, а второй encephale isolee (изолированный передний мозг и изолированный головной мозг). Межколликулярная перерезка оставляет ниже места разреза почти всю ретикулярную формацию ствола; перерезка ниже продолговатого мозга оставляет связи стволовых структур с высшими отделами мозга. У животного с cerveau isolee остаются связанными с корой головного мозга такие важнейшие афферентные системы, как обонятельная и зрительная. Однако такое животное не реагирует на световые и обонятельные раздражения; глаза у него закрыты, и оно фактически не вступает в контакт с внешним миром. У животного же с encephale isolee реакции полностью сохраняются; такое животное открывает глаза на свет, реагирует на запахи, т.е. ведет себя как бодрствующее, а не как спящее животное. Бремер сделал вывод, что для бодрствующего состояния высших отделов мозга необходимо непрерывное поступление к ним афферентной импульсации, в частности от такой обширной рецептивной зоны, как зона тройничного нерва. Предполагалось, что эта импульсация поступает в кору больших полушарий по восходящим афферентным путям и поддерживает высокую возбудимость ее нейронов. Однако дальнейшие исследования показали, что для поддержания бодрствующего состояния коры больших полушарий важно не просто поступление к ней импульсации по афферентным системам. Если сделать перерезку ствола мозга так, чтобы не повредить основные афферентные системы (например, систему медиальной петли), но перерезать восходящие связи ретикулярной формации, то животное тем не менее впадает в сонное состояние, конечный мозг перестает активно функционировать. Следовательно, для поддержания бодрствующего состояния конечного мозга важно, чтобы афферентная импульсация первоначально активировала ретикулярные структуры мозгового ствола. Влияния же из ретикулярных структур по восходящим путям каким то образом определяют функциональное состояние конечного мозга. Проверить такой вывод можно путем прямого раздражения ретикулярных структур. Такое раздражение через погружные электроды было проведено Дж.Моруцци и X.Мегоуном и затем воспроизводилось во многих лабораториях. Оно всегда дает однозначные результаты в виде характерной поведенческой реакции животного. Если животное находится в сонном состоянии, оно просыпается, у него появляется ориентировочная реакция. После прекращения раздражения животное снова возвращается в сонное состояние. Переход от сонного к бодрствующему состоянию в период раздражения ретикулярных структур отчетливо проявляется не только в поведенческих реакциях, он может быть зарегистрирован по объективным критериям деятельности коры больших полушарий, в первую очередь по изменениям ее электрической активности. Для коры больших полушарий характерна постоянная электрическая активность. Ее регистрацию называют электроэнцефалограммой. Активность состоит из небольших по амплитуде (30-100 мкВ) колебаний, которые легко отводятся не только от открытой поверхности мозга, но и от кожи головы. У человека в спокойном дремотном состоянии такие колебания имеют частоту 8-10 в секунду и являются довольно регулярными (альфа-ритм). У высших позвоночных этот ритм менее регулярен, а частота колебаний варьирует от 6-8 у кролика до 15--20 у собаки и обезьяны. Во время активности регулярные колебания сменяются сразу же значительно меньшими по амплитуде и более высокочастотными колебаниями (бета-ритм). Появление периодических больших колебаний ясно говорит о том, что электрическая активность каких-то элементов в коре развивается синхронно. Когда правильные колебания большой амплитуды сменяются низковольтными, частыми колебаниями, то это, очевидно, говорит о том, что клеточные элементы коры начинают функционировать менее синхронно, поэтому такой тип активности называется реакцией десинхронизации. Таким образом, переход от спокойного, неактивного состояния коры к активному связан в электрическом отношении с переходом от синхронизированной активности ее клеток к десинхронизированной. Характерным эффектом восходящих ретикулярных влияний на корковую электрическую активность является именно реакция десинхронизации. Эта реакция закономерно сопровождает описанную выше поведенческую реакцию, характерную для ретикулярных влияний. Реакция десинхронизации не ограничивается каким-нибудь одним участком коры, а регистрируется от больших ее областей. Это говорит о том, что восходящие ретикулярные влияния являются генерализованными. Описанные изменения в электроэнцефалограмме не являются единственным электрическим проявлением восходящих ретикулярных влияний. В определенных условиях можно выявить более прямые эффекты ретикулярной импульсации, поступающей к коре больших полушарий. Они впервые были описаны в 1940 г. американскими исследователями А.Форбсом и Б.Мориссоном, которые исследовали вызванную электрическую активность коры при различных афферентных влияниях. При раздражении какой-либо афферентной системы в соответствующей проекционной зоне коры обнаруживается электрический ответ, указывающий на приход к этой области афферентной волны, этот ответ получил название первичного ответа. Кроме этого локального ответа, афферентное раздражение вызывает длиннолатентный ответ, возникающий в обширных областях коры мозга. Этот ответ Форбс и Мориссон назвали вторичным ответом. То обстоятельство, что вторичные ответы возникают со скрытым периодом, значительно превышающим скрытый период первичного ответа, ясно говорит о том, что они связаны с поступлением афферентной волны в кору не по прямым, а по каким-то окольным связям, через дополнительные синаптические переключения. Позже, когда было применено прямое раздражение ретикулярной формации, было показано, что оно может вызвать ответы такого же типа. Это позволяет заключить, что вторичный ответ является электрическим проявлением поступления в кору больших полушарий афферентной волны через ретикулокортикальные связи. Через ствол мозга проходят прямые афферентные пути, которые после синаптического переключения в таламусе поступают в кору больших полушарий. Приходящая по ним афферентная волна вызывает первичный электрический ответ в соответствующей проекционной зоне с коротким скрытым периодом. Одновременно афферентная волна по коллатералям ответвляется в ретикулярную формацию и активирует ее нейроны. Затем по восходящим путям от нейронов ретикулярной формации импульсация тоже поступает в кору, но уже в виде задержанной реакции, возникающей с большим скрытым периодом. Эта реакция охватывает не только проекционную зону, но и широкие участки коры, вызывая в них изменения, важные для бодрствующего состояния. Дж.Моруцци и сотр. (1941) также обнаружили, что, раздражая некоторые участки ретикулярной формации заднего мозга, можно у животных вызвать в коре вместо десинхронизации синхронизацию электрических колебаний и, соответственно, перевести животное из бодрствующего состояния в пассивное, сонное. Поэтому можно думать, что в составе восходящих путей ретикулярной формации действительно существуют не только активирующие, но и инактивирующие подразделения, последние каким-то образом снижают возбудимость нейронов конечного мозга. Нейронная организация восходящей системы ретикулярной формации не вполне ясна. При разрушении ретикулярных структур среднего и заднего мозга в коре больших полушарий не обнаруживается существенного количества дегенерирующих окончаний, которые могли бы быть отнесены к прямым ретикулярным волокнам. Значительная дегенерация окончаний в коре возникает только при разрушении ядер таламуса, получивших название неспецифических. Поэтому возможно, что восходящие ретикулярные влияния передаются в кору больших полушарий не по прямым путям, а через какие-то промежуточные синаптические соединения, локализованные, вероятно, в промежуточном мозге. Любопытно отметить, что гистологические и электрофизиологические данные указывают на характерную деталь хода аксонов многих ретикулярных нейронов. Аксоны нейронов гигантоклеточного ядра, т. е. основного ядра ретикулярной формации, очень часто делятся Т-образно, и один из отростков идет вниз, образуя ретикулоспинальный путь, а второй вверх, направляясь в верхние отделы головного мозга. Создается такое впечатление, что и восходящие, и нисходящие функции ретикулярной формации могут быть связаны с деятельностью одних и тех же нейронов. По функциональным свойствам ретикулярные структуры, создающие восходящие влияния, также имеют много общего со структурами, обеспечивающими нисходящие влияния. Восходящие влияния несомненно носят тонический характер, они легко изменяются гуморальными факторами и высокочувствительны к фармакологическим веществам. Снотворное и наркотическое действие барбитуратов основано, по-видимому, именно на блокировании в первую очередь восходящих влияний ретикулярной формации. 35.Кора больших полушарий, структурно-функциональная характеристика. Современное представление о локализации функций в коре больших полушарий мозга. Кора головного мозга - это совокупность мозговых отделов, анализаторов. Различные отделы коры головного мозга могут выполнять одновременно и афферентные и эфферентные функции. Мозговой отдел анализатора - состоит из ядра (центральная часть) и рассеянных нервных клеток. Ядро - совокупность высокоразвитых нейронов расположенных в строго определенной зоне коры головного мозга. Поражение ядра приводит к выпадению определенной функции. Ядро зрительного анализатора расположено в затылочной области, мозговой отдел слухового анализатора - в височной области. Рассеянные нервные клетки - менее дифференцированные нейроны, разбросанные по всей коре. В них возникают более примитивные ощущения. Наибольшие скопления этих клеток в теменной области. Эти клетки необходимы, т. к. в них возникают ощущения, которые обеспечивают выполнение функции при поражении ядра. В норме эти клетки обеспечивают связь между различными сенсорными системами. В коре головного мозга существуют проекционные зоны. Первичная проекционная зона - занимает центральную часть ядра мозгового анализатора. Это совокупность наиболее дифференцированных нейронов, в которых происходит высший анализ и синтез информации, там возникают четкие и сложные ощущения. К этим нейронам подходят импульсы по специфическому пути передачи импульсов в коре головного мозга (спиноталамический путь). Вторичная проекционная зона - расположена вокруг первичной, входит в состав ядра мозгового отдела анализатора и получает импульсы от первичной проекционной зоны. Обеспечивает сложное восприятие. При поражении этой зоны возникает сложное нарушение функции. Третичная проекционная зона - ассоциативная - это полимодальные нейроны, разбросанные по всей коре головного мозга. К ним поступают импульсы от ассоциативных ядер таламуса и конвергируют импульсы различной модальности. Обеспечивает связи между различными анализаторами и играют роль в формировании условных рефлексов 36.Вегетативная нервная система - строение, основные физиологические свойства Анатомические и физиологические особенности вегетативной нервной системы Впервые понятие вегетативная нервная система было введено в 1801 г. французским врачом А. Беша. Этот отдел ЦНС обеспечивает экстраорганную и внутриорганную регуляцию функций организма и включает в себя три компонента: 1) симпатический; 2) парасимпатический; 3) метсимпатический. Вегетативная нервная система обладает рядом анатомических и физиологических особенностей, которые определяют механизмы ее работы. Анатомические свойства 1. Трехкомпонентное очаговое расположение нервных центров. Низший уровень симпатического отдела представлен боковыми рогами с VII шейного по III–IV поясничные позвонки, а парасимпатического – крестцовыми сегментами и стволом мозга. Высшие подкорковые центры находятся на границе ядер гипоталамуса (симпатический отдел – задняя группа, а парасимпатический – передняя). Корковый уровень лежит в области шестого—восьмого полей Бродмана (мотосенсорная зона), в которых достигается точечная локализация поступающих нервных импульсов. За счет наличия такой структуры вегетативной нервной системы работа внутренних органов не доходит до порога нашего сознания. 2. Наличие вегетативных ганглиев. В симпатическом отделе они расположены либо по обеим сторонам вдоль позвоночника, либо входят в состав сплетений. Таким образом, дуга имеет короткий преганглионарный и длинный постганглионарный путь. Нейроны пара-симпатического отдела находятся вблизи рабочего органа или в его стенке, поэтому дуга имеет длинный преганглионарный и короткий постганглионарный путь. 3. Эффеторные волокна относятся к группе В и С. Физиологические свойства 1. Особенности функционирования вегетативных ганглиев. Наличие феномена мультипликации (одновременного протекания двух противоположных процессов – дивергенции и конвергенции). Дивергенция – расхождение нервных импульсов от тела одного нейрона на несколько постганглионарных волокон другого. Конвергенция – схождение на теле каждого постганглионарного нейрона импульсов от нескольких преганглионарных. Это обеспечивает надежность передачи информации из ЦНС на рабочий орган. Увеличение продолжительности постсинаптического потенциала, наличие следовой гиперполяризации и синоптической задержки способствуют передаче возбуждения со скоростью 1,5–3,0 м/с. Однако импульсы частично гасятся или полностью блокируются в вегетативных ганглиях. Таким образом они регулируют поток информации из ЦНС. За счет этого свойства их называют вынесенными на периферию нервными центрами, а вегетативную нервную систему – автономной. 2. Особенности нервных волокон. Преганглионарные нервные волокна относятся к группе В и проводят возбуждение со скоростью 3—18 м/с, постганглионарные – к группе С. Они проводят возбуждение со скоростью 0,5–3,0 м/с. Так как эфферентный путь симпатического отдела представлен преганглионарными волокнами, а парасимпатического – постганглионарными, то скорость передачи импульсов выше у парасимпатической нервной системы. Таким образом, вегетативная нервная система функционирует неодинаково, ее работа зависит от особенностей ганглиев и строения волокон. 2. Функции симпатической, парасимпатической и метсимпатической видов нервной системы Симпатическая нервная система осуществляет иннервацию всех органов и тканей (стимулирует работу сердца, увеличивает просвет дыхательных путей, тормозит секреторную, моторную и всасывательную активность желудочно-кишечного тракта и т. д.). Она выполняет гомеостатическую и адаптационно-трофическую функции. Ее гомеостатическая роль заключается в поддержании постоянства внутренней среды организма в активном состоянии, т. е. симпатическая нервная система включается в работу только при физических нагрузках, эмоциональных реакциях, стрессах, болевых воздействий, кровопотерях. Адаптационно-трофическая функция направлена на регуляцию интенсивности обменных процессов. Это обеспечивает приспособление организма к меняющимся условиям среды существования. Таким образом, симпатический отдел начинает действовать в активном состоянии и обеспечивает работу органов и тканей. Парасимпатическая нервная система является антагонистом симпатической и выполняет гомеостатическую и защитную функции, регулирует опорожнение полых органов. Гомеостатическая роль носит восстановительный характер и действует в состоянии покоя. Это проявляется в виде уменьшения частоты и силы сердечных сокращений, стимуляции деятельности желудочно-кишечного тракта при уменьшении уровня глюкозы в крови и т. д. Все защитные рефлексы избавляют организм от чужеродных частиц. Например, кашель очищает горло, чиханье освобождает носовые ходы, рвота приводит к удалению пищи и т. д. Опорожнение полых органов происходит при повышении тонуса гладких мышц, входящих в состав стенки. Это приводит к поступлению нервных импульсов в ЦНС, где они обрабатывают и по эффекторному пути направляются до сфинктеров, вызывая их расслабление. Метсимпатическая нервная система представляет собой совокупность микроганглиев, расположенных в ткани органов. Они состоят из трех видов нервных клеток – афферентных, эфферентных и вставочных, поэтому выполняют следующие функции: 1) обеспечивает внутриорганную иннервацию; 2) являются промежуточным звеном между тканью и экстраорганной нервной системой. При действии слабого раздражителя активируется метсимпатический отдел, и все решается на местном уровне. При поступлении сильных импульсов они передаются через парасимпатический и симпатический отделы к центральным ганглиям, где происходит их обработка. Метсимпатическая нервная система регулирует работу гладких мышц, входящих в состав большинства органов желудочно-кишечного тракта, миокарда, секреторную активность, местные иммунологические реакции и др. 37.Принципы организации эфферентного звена вегетативных рефлексов. Вегетативные ганглии, их функции. Преганглионарные и постганглионарные нервные волокна и их функциональные различия. 38.Вегетативные рефлексы, их классификация. Вегетативные центры. Классификация вегетативных рефлексов: 1)в зависимости от расположения центральных нейронов рефлекса: -истинные: центральный нейрон расположен в ЦНС -местные: центральный нейрон расположен в интрамуральном ганглии -аксон-рефлекс: связан с покраснением кожи при раздражении кожных рецепторов во время массажа, трения; происходит расширение подкожных сосудов 2)в зависимости от места раздражения и места возникновения ответной реакции: -висцеральные т.е. раздражается один внутренний орган, а ответная реакция возникает у другого внутреннего органа(рефлекс Гольца) -висцеросоматические т.е. раздражение внутреннего органа приводит к сокращению мышц туловища -висцеродермальные т.е. раздражение внутреннего органа вызывает изменение кожной чувствительности в определенной области тела(зоны Захарьина-Геда) -дермовисцеральные т.е. раздражение кожных рецепторов приводит к ответной реакции определенного органа Вегетативные центры: 1.низшие: спинной мозг и ствол, в которых расположен центральные нейроны отделов ВНС 2.высшие: -гипоталамус: передний-влияет на парасимпатическую нс, задний-симпатическую -кора больших полушарий, которая приспосабливает вегетативные функции(рефлексы) к меняющимся условиям среды(выработка условных вегетативных рефлексов) 39.Сравнительная характеристика симпатического и парасимпатического отделов вегетативной нервной системы, их влияние на внутренние органы. Сравнительная характеристика симпатического и парасимпатического отделов вегетативной нервной системы
Сравнительная характеристика отдел Признак : Симпатическая нервная система Парасимпатическая нервная система Место расположения Н.Ц.: Грудной и поясничный отдел спинного мозга ПМ,, шейный и поясничный отделы СМ Медиатор: Адреналин,Норадреналин Ацетилхолин(АХ) Влияние на организм: Бодрствование, стресс Система отдыха, восстановления сил Расположение ганглия Рядом со спинным мозгом, в симп.стволе Рядом с рабочим органе либо в его стенках Отличие в рефлекторной дуге: Преганглионарное волокно короткое, постганглионарное длинное(СИМПАТИЧ НС),Преганглионарное волокно длинное, постганглионарное короткое(ПАРА) 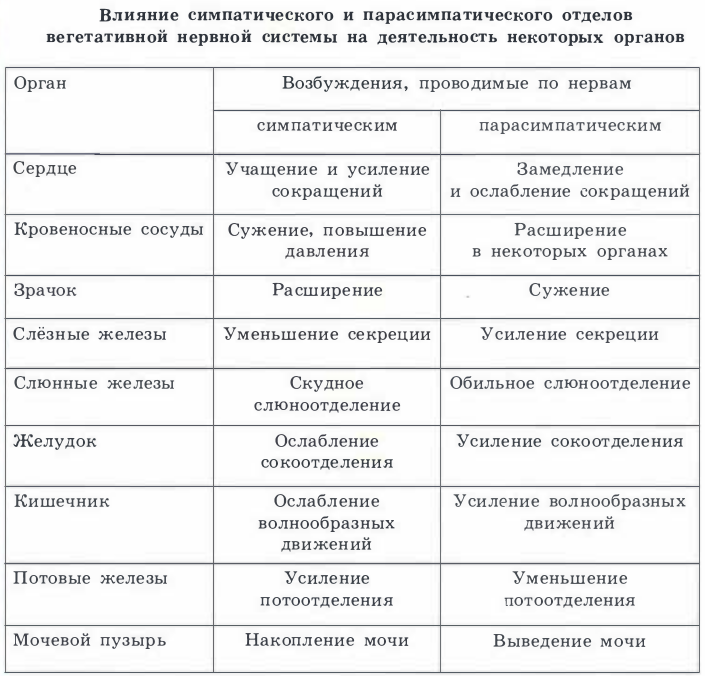 40.Структурно-функциональные особенности соматической и вегетативной нервной системы.(таблица в тетради). 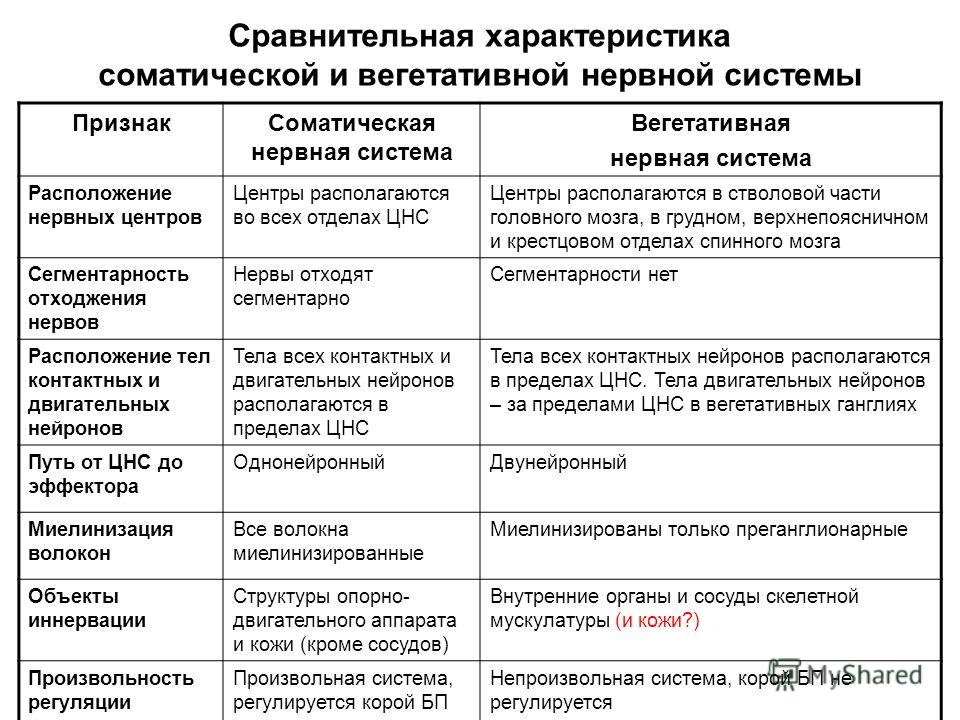 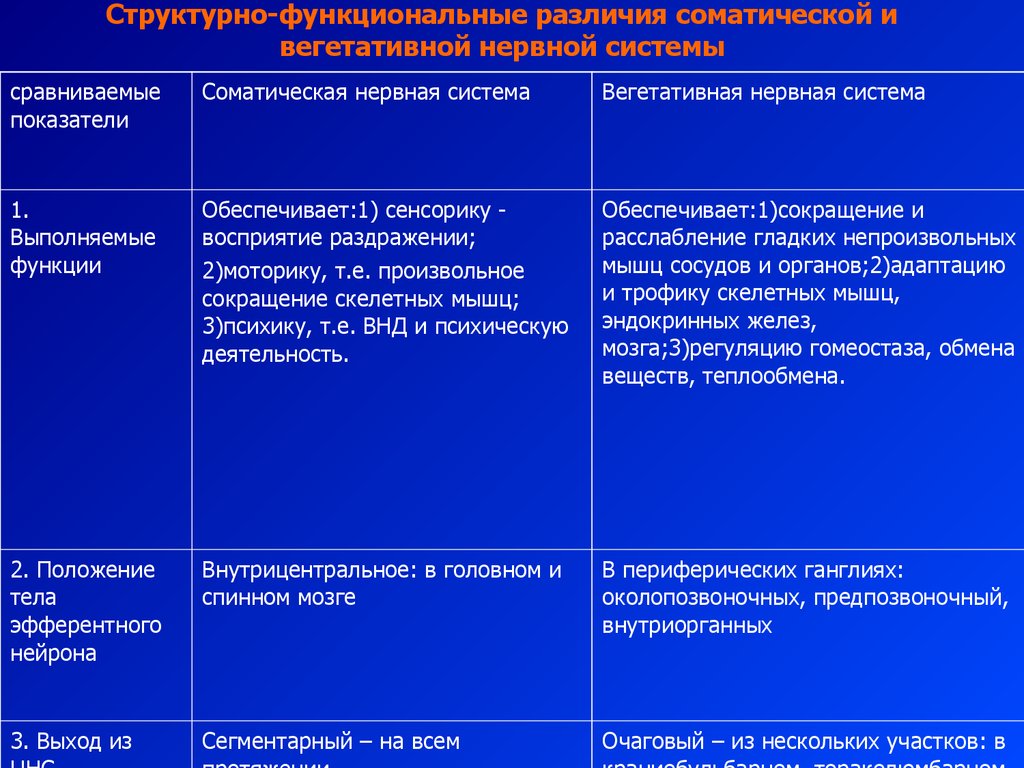 |