Клетка. Занятие 3-4. Клетка. Общая характеристика
 Скачать 1.31 Mb. Скачать 1.31 Mb.
|

2. МикрофиламентыСтроение. Микрофиламенты – самые тонкие компоненты цитоскелета (их толщина 5-7 нм). Обычно они состоят из нескольких типов белка актина. В исчерченных мышечных клетках актиновые филаменты формируют стабильную структуру, связанную с миозиновыми филаментами. Актиновые филаменты в других клетках менее стабильны и могут распадаться (диссоциировать) и повторно собираться. Эти изменения регулируются ионами кальция и циклической АМФ, а также актин-связывающими белками в цитоплазме. Функция. Микрофиламенты обладают сократительной способностью, но для этого они обычно должны взаимодействовать с миозином. В мышечных клетках миозин образует толстые филаменты. В немышечных клетках он существует в растворенном виде. Воздействие цитохолазинами разрушает организацию микрофиламентов и нарушает следующие связанные с ними функции клетки: эндоцитоз, экзоцитоз, сокращение микроворсинок, движения клетки, движения органелл, везикул и гранул, ток цитоплазмы, поддержание формы клетки, разделение дочерних клеток по экватору во время митоза. Локализация. В немышечных клетках микрофиламенты распределены в виде неправильной сети в цитоплазме. Локальные скопления обнаруживаются как тонкая оболочка под плазматической мембраной – терминальная сеть, как параллельные пучки в сердцевине микроворсинок, в цитоплазме у переднего края различных псевдоподий, в ассоциации с другими компонентами цитоплазмы или как ремень (кисет) вокруг экватора делящейся клетки. 3. Промежуточные филаментыСтроение. Промежуточные филаменты, как следует из их названия, занимают промежуточное положение по толщине (10-12 нм) между микротрубочками и микрофиламентами. Они состоят из белков, которые структурно близки с ядерными ламинами и, в отличие от микротрубочек и микрофиламентов, которые устроены одинаково во всех типах клеток, промежуточные филаменты имеют разное строение в различных типах клеток. Белки кодируются примерно 70-ю разными генами. Например, цитокератины характерны для эпителиальных клеток, виментин – для клеток мезенхимного происхождения (фибробластов, хондроцитов), десмин – для мышечных клеток, кислый глиальный фибриллярный белок – для клеток глии, нейрофибриллярный белок – для нервных клеток. Функция белков промежуточных филаментов до конца не изучена. Они, вероятно, участвуют в поддержании формы клетки, возможно, как компонент микротрабекулярной сети. Участвуют в образовании межклеточных контактов (десмосом и полудесмосом)*. Ядерные ламины в настоящее время относятся к V классупромежуточных филаментов, этофибриллярные белки, обеспечивающие структурную функцию и регуляцию транскрипции вядре клеток. *Мутации генов кератинов krt5 и krt14 связаны с развитием наследственного кожного заболевания,буллёзного эпидермолиза(Epidermolysis bullosa simplex). При этом заболевании нарушено прикрепление эпидермиса к базальной пластинке, на коже образуются заполненные серозным содержимым пузыри. Локализация. Большая часть промежуточных филаментов распределены в цитоплазме, вокруг ядра клетки они образуют «корзинку», и от нее распространяются на периферию клетки. III. ЯДРООбщие сведения. Внешние признаки ядер изменяются от ткани к ткани и от клетки к клетке, но каждое ядро имеет ядерную оболочку, хроматин, нуклеоплазму и одно или несколько ядрышек. Хотя некоторые зрелые клетки теряют ядро (например, эритроциты), как минимум одно ядро присутствует на какой-то стадии развития во всех эукариотических клетках. Ядро содержит линейный код (ДНК) для синтеза компонентов клетки и ее продуктов, наделяющих клетку способностью адаптироваться к изменяющимся условиям окружающей среды и внешним сигналам, таким, как гормоны. Микроскопические признаки ядра важны для идентификации и классификации как здоровых, так и поврежденных клеток и тканей. Ядра демонстрируют высокое разнообразие по: 1) размерам, как абсолютным, так и относительным по сравнению с количеством цитоплазмы (ядерно-цитоплазматическое соотношение), 2) количеству на клетку, позволяющему классифицировать клетку как безъядерную, одноядерную, двуядерную или многоядерную, 3) количеству и распределению хроматина и 4) локализации (базальной, центральной, эксцентричной). Ядерная оболочка. Ядерное содержимое отделено от остальной цитоплазмы двойной мембраной, которая называется ядерной оболочкой. Щель между двумя мембранами узкая (40-70 нм), она называется перинуклеарная цистерна, или перинуклеарное пространство. Ядерная оболочка часто рассматривается как выпячивание гранулярной ЭПС, поскольку ее наружная поверхность усыпана рибосомами и имеет связи с гранулярной ЭПС. Внутренняя поверхность внутренней мембраны покрыта фиброзной пластинкой – слоем, состоящим из ядерных белков промежуточных филаментов – ламинов. Ядерная оболочка перфорирована многочисленными отверстиями – ядерными порами, каждая из которых имеет диаметр около 70 нм и окружена 8 глобулярными субединицами, аннулярными белками, которые имеют вид восьмиугольника на некоторых препаратах. Каждая пора покрыта тонкой белковой диафрагмой. Поры образуют каналы для движения важных молекул между ядром и цитоплазмой, в том числе нуклеиновых кислот, синтезируемых в ядре, а используемых в цитоплазме (мРНК, рРНК, тРНК), и белков, синтезируемых в цитоплазме, а используемых в ядре (гистоны, полимеразы). 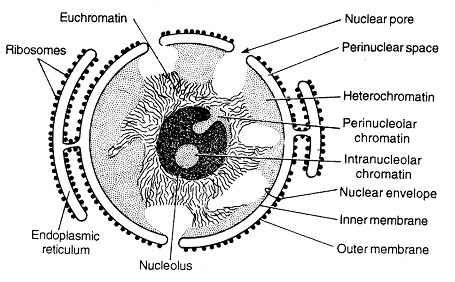 Хроматин – интенсивно базофильный, состоит из ДНК и ассоциированных с ней гистонов и негистоновых белков. Нуклеосомы. Изолированный хроматин виден на электронных микрофотографиях как тонкие нити с выступающими частицами в виде шариков через регулярные промежутки. Каждая нить – это двойная спираль молекулы ДНК, а частицы – повторяющиеся структурные субъединицы хроматина, которые называются нуклеосомами. Каждая нуклеосома состоит из 166 пар оснований нити ДНК, скрученной вокруг сердцевины из 8 гистонов (по 2 копии каждого: Н2А, Н2В, Н3, Н4). Часть нити ДНК между двумя нуклеосомами содержит дополнительно 48 пар оснований и называется линкерным регионом (линкерной ДНК, линкером). Другой гистон (обычно Н1) связан с наружной поверхностью нуклеосомы и линкера. Цепочка из нуклеосом сворачивается в супер-спираль (6 нуклеосом на виток), так образуется конденсированный хроматин – гетерохроматин. 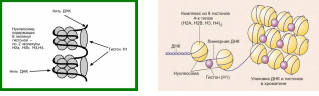 Типы хроматина. Ядро содержит высоко конденсированный хроматин (гетерохроматин), который окрашивается в темный цвет основными красителями. Поскольку для транскрипции с ДНК она должна быть раскручена, 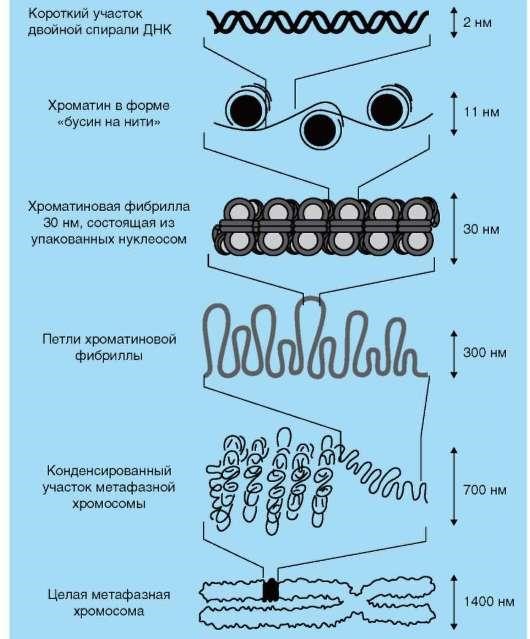 клетки с темно окрашенными ядрами (содержащими гетерохроматин) менее активны в транскрипции ДНК и используют меньшую часть их общего генома по сравнению с другими клетками. Несвернутый хроматин (эухроматин) окрашивается плохо и его трудно различить даже в электронный микроскоп. Большие, бледно окрашенные ядра (эухроматин) обычно бывают у клеток с высокой активностью транскрипции и быстрыми клеточными делениями. Паттерны хроматина. Количество и распределение ядерного хроматина часто импользуют для идентификации клеточного типа, особенно для клеток, которые не имеют характерного окрашивания цитоплазмы. Даже в большинстве эухроматических ядер ободок гетерохроматина часто обнаруживается на внутренней поверхности ядерной оболочки в ассоциации с фиброзной пластинкой. Этот связанный с ядерной оболочкой гетерохроматин позволяет увидеть границы ядра в световой микроскоп. Хромосомы, наиболее конденсированная форма хроматина, становятся видны во время митоза. Супер-спирали гетерохроматина образуют складки и накручиваются на центральную основу из негистоновых белков. В клетках человека имеется 46 хромосом, 44 соматических хромосомы образуют 22 пары структурно похожих хромосом. Одна пара половых хромосом состоит из двух различных (XY) хромосом у мужчин и двух похожих (XX) хромосом у женщин. У женщин используется только одна из пары Х-хромосом. Неактивная Х-хромосома часто видна как глыбка гетерохроматина, которую называют половой хроматин, или тельце Барра. В большинстве клеток тельце Барра находится у внутренней поверхности ядерной оболочки. В нейрофилах оно может появиться как отросток от сегментированного ядра в виде барабанной палочки. Кариотип. Кариотип клетки – это перечень ее хромосом или изображение ее хромосом, разложенных по типам. Приготовление такого изображения называется кариотипированием. Клетки в культуре стимулируют к митозу фитогемагглютинином (митоген растительного происхождения). Делящиеся клетки обрабатывают колхицином (вспомните механизм и точку приложения действия колхицина), чтобы остановить митоз в метафазе, когда хромосомы высоко конденсированы и хорошо видимы. Лизис клеток с помощью гипотонического раствора позволяет хромосомам распределиться на стекле с минимальным наложением друг на друга или без такого наложения. Далее хромосомы фотографируют, собирают в пары и располагают в определенной последовательности. Кариотипирование позволяет каталогизировать хромосомы для выявления их структурных нарушений, потери или появления избыточных хромосом. Ядрышко. В ходе интерфазы (между митозами) каждое ядро обычно имеет как минимум одно интенсивно базофильное тело, которое называется ядрышком. Ядрышки – места синтеза большей части рРНК. Обычно они отличаются от гетерохроматина, но в темных ядрах могут быть видны плохо. Наиболее многочисленные и крупные ядрышки встречаются в эмбриональных клетках, в клетках, активно синтезирующих белки, и в быстро растущих злокачественных опухолевых клетках. Небольшие количества гетерохроматина прикрепляются к ядрышкам. Значение такого ядрышкоассоциированного хроматина неизвестно. При подготовке клетки к митозу ядрышки исчезают и снова появляются после завершения митоза. Под электронным микроскопом можно увидеть различные компоненты ядрышка. Аморфная часть – бледно окрашенная часть ядрышка, содержащая ядрышковый организатор ДНК, который несет код для рРНК. У человека 5 пар хромосом имеют имеют регионы ядрышкового организатора. Таким образом, возможно образование 10 ядрышек, но слияние ядрышковых организаторов приводит к появлению меньшего числа более крупных ядрышек. Новая синтезированная рРНК появляется первой в этом регионе. Нуклеолонема – термин, который используют для обозначения нитевидной базофильной структуры ядрышка. Нуклеонема содержит 2 компонента, богатых рРНК и различимых в ЭМ. Фиброзная часть состоит из плотно упакованных рибонуклеобелковых волокон, 5-10 нм в диаметре. Эти волокна состоят из вновь синтезированных первичных транскриптов генов рРНК и ассоциированных с ними белков. Вновь синтезированная рРНК появляется второй в этом регионе. Гранулярная часть, 15-20 нм в диаметре, которые представляют собой созревающие рибосомальные субъединицы в процессе сборки для экспорта в цитоплазму. Вновь синтезированная рРНК появляется третьей в этом регионе. Нуклеоплазма. Это матрикс, в котором находятся другие внутриядерные компоненты. Он состоит из ферментных и неферментных белков, метаболитов, ионов и воды. Он включает ядерный матрикс – фибриллярную ядерно-скелетную структуру, которая появляется, чтобы связывать некоторые рецепторы гормонов и вновь синтезированную ДНК. 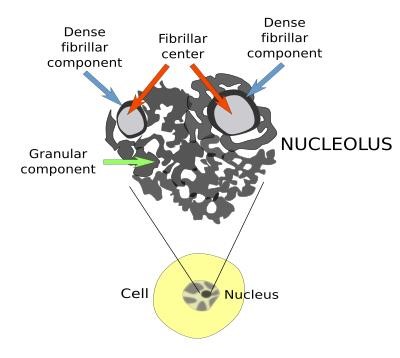 |