Пьер Тейяр де Шарден. Феномен человека. Пьер Тейяр де Шарден феномен человека перевод и примечания Н. А. Садовского М. "Прогресс", 1965 к читателю
 Скачать 1.18 Mb. Скачать 1.18 Mb.
|

3. ДРЕВО ЖИЗНИА. Основные черты а. Одна из количественных единиц эволюции – пласт млекопитающих Из предшествующих замечаний непосредственно вытекает, что для четкого представления древа жизни необходимо "натренировать взгляд" на той части его кроны, на которой лишь умеренно отразилось разрушающее действие времени. Ни слишком близко, чтобы листья не мешали видеть; ни слишком далеко, чтобы ветви были еще достаточно густыми. Где найти в нынешней природе этот привилегированный участок? Безусловно, в большой семье млекопитающих. Мы положительно знаем из геологии (достаточно обратить внимание на ее внутреннюю структуру), что хотя человечество еще "незрелая" группа, в целом млекопитающие – группа взрослая и в то же время свежая. Они полностью развились лишь в третичном периоде, и среди них до сих пор сохранилось значительное количество его самых деликатных остатков. Вот почему эта группа с самого начала была и остается излюбленным объектом при возникновении и развитии трансформистских идей. Рассмотрим эту группу в ее основных чертах (рис. 1). Однако для начала ограничим поле наших исследований самой молодой и прогрессивной частью этой группы: плацентарными млекопитающими *) *) Так называют в противоположность аплацентарным (или сумчатым) млекопитающих, у которых зародыш благодаря наличию специальной оболочки – плаценты, которая его защищает и питает, может до полного созревания оставаться в утробе матери. С эволюционной (можно даже сказать, с "физиологической") точки зрения плацентарные млекопитающие, взятые в целом, составляют то, что я условно здесь называю биот. Под этим я подразумеваю мутовочную группировку, элементы которой не только родственны по рождению, но и, кроме того, поддерживают и взаимно дополняют друг друга в борьбе за существование и распространение. Чтобы понять этот важный пункт, выясненный главным образом американской школой палеонтологии,[11] достаточно рассмотреть в соответствующем свете распределение наиболее знакомых каждому из нас животных форм. Здесь – травоядные и грызуны, извлекающие свою пищу непосредственно из растительного ответвления жизни; там – насекомоядные, паразитирующие подобным же образом на "членистоногих". Здесь – плотоядные, существующие за счет тех и других, а там – всеядные, питающиеся за всеми столами сразу. Таковы четыре главные радиации, в основном совпадающие с общепринятым делением фил. Рассмотрим теперь эти четыре луча или сектора один за другим, раздельно. Каждый из них совершенно свободно подразделяется, расщепляется на меньшие подразделения. Возьмем, например, ту ветвь, которая сейчас представляется самой густой, – травоядных. В зависимости от двух различных способов, избранных для превращения конечностей в бегающие лапы (путем исключительного развития двух пальцев или одного среднего пальца), появляются два крупных семейства – парнокопытные и непарнокопытные, каждое из которых образуется из пучка отдельных больших линий. Здесь у непарнокопытных – серая толпа тапирообразных, короткая, но удивительная ветвь титанотериев, халикотерии с землероющими когтями, которых человек, вероятно, еще застал, племя носороговых, безрогих или с рогом, и, наконец, однокопытные лошади с параллельной им совершенно независимой филой в Южной Америке. Там, у парнокопытных – свиньи, верблюды, олени и антилопы, не говоря уже о других стеблях, менее жизнеустойчивых, но столь же индивидуализированных и интересных с точки зрения палеонтологии. А ведь есть еще обширная и мощная группа хоботных! Согласно правилу "устранения черешков", начало каждого из этих подразделений тонет в тумане прошлого. Но раз уж они появились, то мы можем проследить, у всех вместе и у каждого в отдельности главные фазы их географического распространения; их последовательное деление, почти до бесконечности, на субмутовки; наконец, чрезмерное усиление путем ортогенеза некоторых особенностей костей, зубов или черепа, что обычно превращает животных в уродов или делает их недолговечными. Но и это еще не все. Поверх этого обилия родов и видов, образовавшихся из четырех основных радиации, мы различаем другую сеть, соответствующую сделанным там и сям попыткам покинуть наземную жизнь и оккупировать воздух, воду и даже глубины земли. Наряду с формами, приспособленными для бега, появляются лазающие и летающие, плавающие и землероющие. 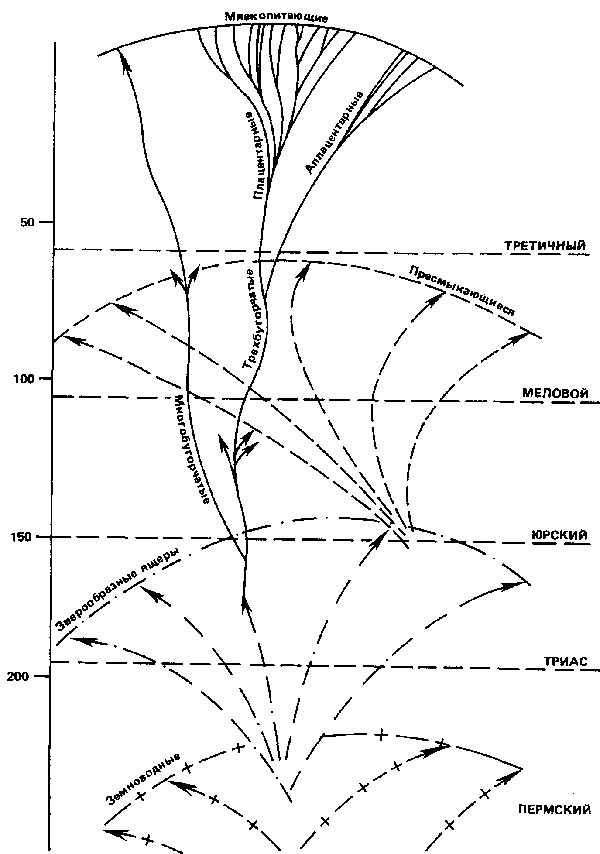 Рис. 1. Схема, символизирующая развитие слоями четвероногих. Цифры слева обозначают миллионы лет. Детальное объяснение в тексте. Одни (киты и сирены), очевидно, с удивительной быстротой произошли от хищных и травоядных. Другие (рукокрылые, кроты и земляные крысы) – главным образом от наиболее древних элементов плацентарной группы – насекомоядных и грызунов, существующих с конца вторичного периода. Рассматривая этот столь хорошо сбалансированный функциональный ансамбль сам по себе, нельзя не признать, что он представляет собой органическую естественную группировку sui generis. Это убеждение еще более крепнет, когда замечаешь, что здесь не исключительный и изолированный случай, а что подобные единства складывались в ходе истории жизни неоднократно. Приведем только два примера, опять-таки не выходя за пределы млекопитающих. В третичный период, учит геология, часть плацентарного биота, находившегося тогда в состоянии глубокой эволюции, оказалась отрезанной морем в южной половине Американского континента. Как же реагировал этот черенок на свою изоляцию? Точно так же, как реагирует черенок растения, то есть воспроизводя в меньшем масштабе рисунок ствола, от которого был отделен. Он начал выпускать отростки своих псевдохоботных, своих псевдогрызунов, своих псевдолошадей, своих псевдообезьян (широконосые обезьяны)... Целый биот в уменьшенном виде (подбиот) внутри первоначального биота! Теперь второй пример, касающийся сумчатых. Судя по их сравнительно примитивной форме размножения, а также по их современному географическому распространению, явно несплошному и остаточному, сумчатые (аплацентарные) представляют собой особый ярус у основания млекопитающих. Они, очевидно, достигли расцвета раньше плацентарных и образовали раньше их свой собственный биот. В целом, за исключением нескольких странных типов (вроде псевдо-махайрода, ископаемые останки которого недавно найдены в Патагонии) *), этот биот сумчатных бесследно исчез. Но зато один из его подбиотов, развившийся еще до третичного периода и случайно сохранившийся опять же вследствие изоляции в Австралии, до сих пор вызывает восхищение натуралистов четкостью своих контуров и своим совершенством. Как всем известно, Австралия при открытии ее европейцами была населена лишь сумчатыми **), но зато сумчатыми всех размеров, всех зон обитания, всех форм – сумчатыми травоядными и бегающими, хищными сумчатыми, насекомоядными сумчатыми, сумчатыми крысами, сумчатыми кротами и т. д. Нельзя себе представить более яркий пример способности, присущей каждой филе, к дифференциации в своего рода замкнутый, физиологически полный организм. *) Махайрод, или саблезубый тигр. Эта крупная кошка, весьма распространенная в конце третичного и начале четвертичного периодов, имеет странный дубликат в лице жившего в плиоцене хищного сумчатого Южной Америки. **) Кроме одной группы грызунов и прибывших последними человека и его собаки. Установив это, подымимся выше, чтобы рассмотреть обширную систему двух биотов – плацентарного и аплацентарного, вместе взятых. Зоологи очень быстро заметили, что у всех форм, входящих в эти две группы, коренные зубы состоят в основном из трех бугорков, зацепляющихся от одной челюсти к другой, сверху вниз. Черта, незначительная сама по себе, но тем более интригующая своим постоянством. Как объяснить универсальность столь случайной черты? Ключ к отгадке был дан открытием, сделанным в некоторых юрских отложениях в Англии. В средней юре на какое-то мгновение нам приоткрывается первая пульсация млекопитающих – мир маленьких зверьков, не больше крыс или землероек. И вот у этих-то мелких, но уже чрезвычайно разнообразных животных тип зубов еще не закреплен, как в нынешней природе. Среди них уже можно различить и трехбугорчатый тип. Но наряду с ним в развитии и способах противоположения бугорков на коренных зубах можно наблюдать и всякие другие комбинации. И эти другие комбинации уже давно устранены! Напрашивается вывод. За исключением, может быть, утконоса и ехидны (этих яйценосных парадоксальных форм, в которых предполагали продолжение многобугорчатых), все ныне живущие млекопитающие ведут начало от одного узкого пучка. Все вместе они представляют собой (в развитом состоянии) лишь один из многочисленных побегов, на которые делится юрская мутовка млекопитающих – побег трехбугорчатых... *) *)...которых можно также назвать "семипозвонковыми", потому что по столь же неожиданному и столь же замечательному совпадению все они имеют семь шейных позвонков, какова бы ни была длина их шеи. Здесь мы почти достигли предела того, что можно видеть сквозь толщу прошлого. Ниже, кроме вероятного существования в самом конце триаса еще одной мутовки, к которой относятся многобугорчатые, история млекопитающих теряется во мраке. По крайней мере около этого времени и выше их группа, естественно изолированная из-за отрыва черешка, выделяется с достаточной четкостью и индивидуальностью, чтобы мы могли ее взять в качестве практической единицы "эволюционной массы". Назовем эту единицу пластом. Нам тут же придется ею пользоваться. б. Пласт пластов – четвероногие Когда необходимо измерить расстояние до туманностей, астрономы пользуются световыми годами. В свою очередь, если мы хотим, отправляясь от млекопитающих, расширить и продолжить вниз наше видение древа жизни, нам надо отсчитывать пластами. Для начала возьмем пласты пресмыкающихся вторичного периода. Когда ниже юрского периода мы теряем из виду ветвь млекопитающих, то она не рассеивается в какой-то пустоте. Ее покрывает и обволакивает густая листва жизни совершенно иного вида: динозавры, птерозавры, ихтиозавры, крокодилы и масса других чудищ, менее знакомых для людей, не сведущих в палеонтологии. В этом ансамбле зоологические расстояния между формами явно более велики, чем между отрядами млекопитающих. Однако три особенности бросаются в глаза. Прежде всего здесь мы имеем дело с разветвленной системой. В этой системе, далее, ветви выступают на уже зрелой или даже конечной стадии распускания. Наконец, вся группа в целом представляет собой не что иное, как огромный и, может быть, сложный биот. Тут – травоядные, зачастую гигантских размеров. Там – их спутники и тираны – плотоядные, массивные или прыгающие. Здесь – летающие, с перепонками летучих мышей или перьями птиц. И, наконец, плавающие, такие же обтекаемые, как и дельфины. На расстоянии этот мир пресмыкающихся нам представляется более сжатым, чем мир млекопитающих, но его длительность, измеряемая конечной экспансией и усложнением, может считаться по крайней мере равной. Во всяком случае, он также исчезает в глубинах времен. К середине триасадинозавров еще можно заметить. Но они едва выступают из другого пласта, почти пришедшего к своему закату, – пласта пермских пресмыкающихся, для которых особенно характерны зверообразные ящеры. Толстые и бесформенные, к тому же редкие в наших музеях, зверообразные значительно менее популярны, чем диплодоки и игуанодоны. Но это не мешает им приобретать все большее значение в зоологии. Если раньше на них смотрели как на странные, диковинные существа, узко ограниченные пределами Южной Африки, то теперь окончательно признано, что они одни представляют целую стадию жизни континентальных позвоночных. В определенный период, до динозавров, до млекопитающих, именно они занимали всю не покрытую морем территорию и владели ею. Или вернее будет сказать, что эти ящеры, крепко стоявшие на своих хорошо сочлененных конечностях, зачастую обладавшие "корневидными" зубами, были первыми четвероногими, прочно утвердившимися на суше. В тот момент, когда мы их обнаруживаем, они уже изобилуют в самых причудливых формах – с рогами, с гребнями, с бивнями, что (как всегда!) указывает на группу, достигшую конца своей эволюции. Фактически же при всей своей внешней причудливости это довольно монотонная группа, в которой еще не различаются ясно прожилки настоящего биота. И все же эта группа поражает развертыванием вширь и потенциальными возможностями своей мутовки. С одной стороны, неподвижные черепахи. С другой – чрезвычайно прогрессивные по своей подвижности и строению черепа типы, из которых, как мы имеем все основания предполагать, и выступил давно дремавший стебель млекопитающих. А дальше новый "провал". На этих расстояниях, под тяжестью прошлого отрезки длительности быстро сжимаются. Когда у основания пермского периода и ниже его мы снова различаем поверхность обитаемой Земли, то она оказывается заселенной лишь земноводными, ползающими по тине. Земноводные – изобилие приземистых или змеевидных тел, среди которых зачастую трудно отличить личинки от взрослых особей; голая или покрытая чешуей кожа; трубчатые или в виде мозаики косточек позвонки... И здесь, согласно общему правилу, нам удается схватить уже только высокодифференцированный мир, почти пришедший к своему концу. Сколько еще пластов мы, может быть, смешиваем в этом кишении, исследуя осадки, толщину и громадность истории которых мы еще плохо осознаем. Но по крайней мере достоверно одно – на этой стадии мы схватываем животную группу, которая начинает выходить из воды-кормилицы, в которой она образовалась. Но уже в этом самом начале своей субвоздушной жизни позвоночные демонстрируют перед нами одну удивительную особенность, над которой следует поразмыслить. У всех у них "одна и та же структура скелета; в особенности она тождественна (не говоря уже о поразительных соответствиях черепа) в отношении числа и плана двигательных конечностей. В чем причина этой схожести? То, что все земноводные, пресмыкающиеся и млекопитающие имеют четыре, и только четыре, лапы, может в крайнем случае объясняться просто стремлением к наиболее простой форме передвижения (однако насекомые никогда не имеют менее шести ног...). Но как объяснить чисто механическими причинами совершенно одинаковую структуру этих четырех отростков? Впереди – единственная плечевая кость, за ней две кости предплечья, затем пять лучей руки?.. Не представляет ли это еще раз одну из тех случайных комбинаций, которые были открыты и реализованы лишь однажды? Здесь снова поневоле напрашивается вывод, уже сделанный в случае трехбугорчатости млекопитающих. Несмотря на свое чрезвычайное разнеобразие, обладающие легкими наземные животные представляют собой не что иное, как вариации, нагроможденные на совершенно частном решении жизни. Таким образом, огромный и сложный веер ходячих позвоночных, будучи продолжен к своим истокам, снова свертывается и смыкается в один луч. Один-единственный черешок замыкает и определяет у его основания пласт пластов – мир четвероногих. в. Ветвь позвоночных В случае млекопитающих мы сумели схватить мутовку, из которой выделился и устремился вперед побег "трехбугорчатых". Что же касается происхождения земноводных, то тут наука не достигла таких успехов. Однако можно без колебаний указать ту единственную область жизни, в которой среди других испробованных комбинаций могли образоваться четвероногие. Они должны были возникнуть где-то среди рыб с дольчатыми и "членообразными" плавниками, некогда жизнеустойчивый пласт которых ныне существует лишь в виде нескольких живых ископаемых: двоякодышащих (или рыб, обладающих легкими) и – совсем свежий сюрприз – кистеперой рыбы (Crossopterigien), недавно выловленной в южных морях. Поверхностно "гомогенизированные" путем механической адаптации к плаванию, рыбы (лучше сказать рыбообразные) невероятно сложны в своей совокупности. Сколько пластов, особенно здесь, соединено и смешано под одним и тем же названием?.. Сравнительно молодых пластов, развившихся в океане в ту же эпоху, когда на континентах распространились пласты четвероногих. И еще более многочисленных старых пластов, заканчивающихся очень низко, у силура, основной мутовкой, от которой, на наш взгляд, расходятся два главных побега: бесчелюстные рыбообразные, с единственной ноздрей, представленные в нынешней природе одной лишь миногой; и челюстные рыбообразные, с двумя ноздрями, от которых пошло все остальное. После всего вышесказанного о последовательном развитии земных форм я не буду пытаться затрагивать и расчленять этот другой мир. Я постараюсь лучше обратить внимание на факт другого порядка, с которым мы встречаемся здесь впервые. Самые древние рыбы, которых мы знаем, в большинстве случаев облечены в мощный, даже чересчур мощный панцирь. *) Но под этой первой, очевидно довольно безуспешной, попыткой упрочения извне скрывается скелет еще целиком из хрящей. По мере следования за ними вниз позвоночные выступают перед нами все менее окостенелыми внутри. И этим объясняется тот факт, что даже в неповрежденных отложениях мы полностью теряем их следы. Но в данном частном случае проявляется общее явление, имеющее важнейшее значение. Какую бы живую группу мы ни рассматривали, в глубине она всегда тонет в сфере мягкого. Безошибочный способ скрыть свой черешок... *) Без этого окостеневшего покрова от них ничего бы и не осталось, и мы бы о них ничего не знали. Ниже девонского периода рыбообразные вступают в своего рода фетальную или личиночную фазу, не поддающуюся окаменению. Не доживи случайно до нас странный ланцетник, мы бы не имели никакого понятия о многочисленных этажах, из которых, по-видимому, строился тип хордовых, до тех пор пока он стал в состоянии заполнить воду, а в последующем и сушу. Так заканчивается и ограничивается у основания большой пустотой громадное сооружение всех четвероногих и всех рыб – ветвь позвоночных. г. Остальная жизнь Ветвь – это наиболее широкий тип оформленной совокупности, еще признаваемый систематикой внутри биосферы. Помимо позвоночных, основную крону жизни образуют еще только две ветви – ветвь червей и членистоногих и ветвь растений. Приобретая прочность – одна с помощью хитина и известняка, другая с помощью целлюлозы, – они также сумели вырваться из плена воды и широко распространиться в воздушной среде. В результате растения и насекомые в нынешней природе перемешались и борются с костистыми животными за то, кто больше займет места в мире. Для каждой из этих двух ветвей можно было бы повторить анализ (но я не стану этого делать), предпринятый в предыдущих параграфах о позвоночных. Вверху – молодые группы, богатые легкими мутовками. Глубже – пласты с более четко выраженными, но менее густыми разветвлениями. В самом низу – погружение в мир химически неустойчивых форм. Та же самая общая картина развития. Но, так как в данном случае ветви, конечно, старше, то здесь и большее усложнение; а в случае насекомых – крайние формы социализации. Не может быть сомнения, что в бездне времен эти различные линии сходятся к какому-то общему полюсу рассеивания. Но задолго до того, как смыкаются хордовые, кольчатые черви и растения (две первые ветви, очевидно, среди многоклеточных; эти последние и растения – только на уровне одноклеточных существ), их стволы исчезают в скоплении причудливых форм – губок, иглокожих, полипов... Все это – наброски ответов на проблему жизни. Пучок неудавшихся ветвей. Все они, несомненно, возникли (хотя мы и не можем сказать, каким образом, – столь глубоким из-за действия длительности стал разрыв) из другого, невероятно древнего и многообразного мира – инфузорий, различных простейших, бактерий, свободных клеток, обнаженных или покрытых щитком, у которых царства жизни смешались, и систематика бессильна. Животные это или растения? Эти слова более не имеют смысла. Наслаивание пластов и ветвей, или "мицелий", перепутанных волокон, как у гриба? Этого мы уже не знаем. Не знаем мы также, из чего это все зародилось. Начиная с докембрия и одноклеточные в свою очередь теряют свой остов из кремния или известняка. И тем самым в мягкости тканей и в метаморфозе первородного ила окончательно исчезают с наших глаз корни древа жизни. Б. Размеры Вот и набросана, хотя и весьма схематично, структурная картина форм, собранных и классифицированных упорным трудом натуралистов, начиная с Аристотеля и Линнея. В ходе описания мы уже старались дать почувствовать огромную сложность этого возрожденного нами мира. Перед лицом всей совокупности нам надо, однако, последним усилием видения более ясно осознать эту громадность размеров. Сам по себе наш рассудок постоянно склонен не только прояснять (что составляет его функцию), но и сужать и укорачивать реальности, к которым прикасается. Устав, он сгибается под тяжестью расстояний и множеств. Так что, обрисовав с грехом пополам экспансию жизни, нам теперь надо представить себе элементы нашей схемы в их истинных размерах – как по численности, так и по объему и по длительности. Попытаемся это сделать. Прежде всего по численности. Для простоты наш эскиз живого мира делался с помощью широких коллективных групп – семейств, отрядов, биотов, пластов, ветвей... Но. пользуясь этими различными единицами, думали ли мы о множествах, с которыми реально имели дело? Пусть тот, кто хочет осмыслить или описать эволюцию, прежде всего отправится побродить в один из тех четырех-пяти крупнейших музеев мира, в которых (ценой усилий, героизм и духовное значение которых будут однажды оценены) легион путешественников сумел в сжатом виде в нескольких залах представить весь спектр жизни. И пусть он там, не обращая внимания на названия, как следует проникнется тем, что его окружает. Тут – мир насекомых, где полноценные виды исчисляются десятками тысяч. Там – моллюски, другие тысячи, бесконечно разнообразные по рисунку прожилок и форме завитков. Затем идут рыбы, с такими же неожиданными, капризными и пестрыми формами, как у бабочек. Далее птицы, едва ли менее фантастические, самого разного профиля, со всякими клювами, всех расцветок. Потом антилопы, всех мастей, с разными осанками, со всевозможными диадемами и т. д. и т. д. Какое многообразие, какой, порыв, какое бурление под каждым из этих названий, которые вызывали в нашем воображении какую-нибудь дюжину весьма привычных форм! А ведь перед нами здесь только ныне живущие. Что, если бы мы могли видеть всех остальных... Во все эпохи Земли, на всех стадиях эволюции другие музеи зарегистрировали бы такое же бурление, такое же буйное изобилие. Все эти сотни тысяч названий, внесенных в каталоги нашей систематики, не представляют и миллионной доли листьев, произраставших до сего дня на древе жизни. Теперь по объему. То есть каково количественное соотношение между различными зоологическими и ботаническими группами в природе? Какова материально доля каждой из них в общей совокупности организованных существ? Чтобы дать общее представление об этой пропорции, я воспроизвожу здесь (рис. 2) выразительный рисунок, которым один ученый натуралист г. Кено представил, по самым последним данным науки, карту животного царства с его основными подразделениями. Эта карта больше позиционная, чем структурная, но она точно отвечает на поставленный вопрос.  Рис. 2. Древо жизни, по Кено (изд. Массон и К°). На этом символическом рисунке каждая главная долька (или гроздь) соответствует одному пласту, по крайней мере столь же важному (морфологически и количественно), как пласт, составляемый всеми млекопитающими. Ниже линии АВ – формы, живущие в воде; выше ее – живущие в воздушной среде. Рассмотрим эту схему[12]. Не правда ли, при первом же взгляде наш рассудок испытывает шок, подобно тому шоку, который мы чувствуем, когда астроном показывает Солнечную систему как простую звезду, все наши звезды – как один Млечный Путь, а наш Млечный Путь – как атом среди других Галактик?.. Млекопитающие, в которых обычно воплощается для нас идея и образ "животного", жалкая долька, поздно возникшая на стволе жизни! А зато что делается вокруг них! А рядом? А ниже?.. Какое обилие соперничающих типов, о существовании, величине, множестве которых мы и не подозревали! Таинственные существа, которых мы могли случайно видеть скачущими среди сухих листьев или ползающими по песку, ни разу не задаваясь вопросом, что они означают и откуда взялись. Существа, незначительные по своим размерам и ныне, быть может, по числу... Эти оставленные в пренебрежении формы теперь предстают в своем истинном свете. По богатству своих разновидностей, по времени, которое потребовалось природе для их порождения, каждая из них представляет собой столь же важный мир, как и наш. Количественно (я подчеркиваю это) мы – лишь одни из них и притом возникшие последними. Наконец, по длительности. Здесь, как обычно, нашему воображению трудно восстановить подлинную картину. Как я уже отмечал, еще более неумолимо, чем горизонты пространства, в нашей перспективе сближаются и налезают друг на друга плоскости прошлого. Как же их разделить? Чтобы придать глубинам жизни их истинный рельеф, полезно для начала вернуться к тому, что я выше назвал пластом млекопитающих. Поскольку этот пласт относительно молод, у нас есть некоторое представление о времени, потребовавшемся для его развития начиная с момента, когда он явственно поднялся над пресмыкающимися в конце мелового периода. Весь третичный период и еще немного. Каких-нибудь 80 миллионов лет. Допустим теперь, что на оси одной и той же зоологической ветви пласты образуются периодически, как сучки вдоль ствола хвойного дерева; так что максимумы их расцвета (единственно четко регистрируемые) в случае позвоночных следуют друг за другом на расстоянии в 80 миллионов лет. Чтобы получить величину длительности одного зоологического интервала, достаточно помножить 80 миллионов лет на число пластов в данном интервале. Так, между млекопитающими и началом четвероногих имеется не менее трех пластов. Числа становятся внушительными. Но они достаточно хорошо согласуются с представлениями, которые вырабатывает геология, о громадности триасового, пермского и каменноугольного периодов. Можно применить и другой, более приблизительный метод, основанный на сравнении ветвей. Внутри одного пласта (возьмем снова пласт млекопитающих) мы в состоянии примерно определить среднее отклонение форм между собой – для всего этого рассеивания потребовалось, повторяем, около 80 миллионов лет. После этого сравним между собой млекопитающих, насекомых и высшие растения. Если три ветви, на конце которых цветут эти три группы, расходятся из одного и того же корня, а не возникли раздельно на одном "мицелии" (что возможно), то какое же требовалось время, сколько периодов, чтобы создать эти гигантские разрывы между типами! Здесь уже зоология бросает вызов данным, приводимым геологией. Прошло всего лишь 1 500 миллионов лет со времени самых древних следов углерода в осадках, решили физики, измерив процент содержания свинца в одном радиоактивном минерале докембрия. Но не предшествуют ли первые организмы этим первым следам? И потом в случае разногласия при подсчете возраста Земли – на какой из двух хронометров следует положиться? На медленность разложения радия? Или на медленность "сложения" живой материи? Если простой секвойе для своего полного роста требуется пять тысяч лет (и еще никто не видел, чтобы секвойя погибла своей естественной смертью), то каков же должен быть полный возраст древа жизни. В. Очевидность И вот теперь это дерево перед нами. Странное это дерево. Можно сказать, негатив дерева, ибо в отличие от гигантов наших лесов его ветви, его ствол обнаруживают себя перед нами лишь пустотами все большего диаметра. С виду – застывшее дерево, столь длительным кажется нам раскрытие почек, которые мы всегда будем узнавать лишь полураскрытыми. Но тем не менее дерево, ясно очерченное своей многоярусной листвой наблюдаемых видов. В своих основных чертах и размерах оно стоит перед нами, покрывая Землю. Рассмотрим его получше, прежде чем проникать в секрет его жизни. Ибо уже из простого созерцания его внешних форм можно извлечь один важный урок – чувство его очевидности. В мире еще встречаются люди, подозрительно и скептически настроенные в отношении эволюции. Зная природу и натуралистов лишь по книгам, они полагают, что борьба вокруг трансформизма все еще продолжается, как во времена Дарвина. А поскольку в биологии идут дискуссии относительно механизма видообразования, они воображают, что эта наука сомневается или еще могла бы сомневаться, не отрицая самой себя, насчет факта и реальности такого развития. Но положение совершенно иное. Может показаться удивительным, что в этой главе, посвященной последовательной цепи развития организованного мира, я ни разу не упомянул о продолжающихся оживленных спорах по вопросу о различии между сомой и зародышевым веществом, о существовании и функциях генов, о передаваемости или непередаваемости приобретенных признаков. Но это потому, что на данном этапе моего исследования эти вопросы меня непосредственно не интересуют. Для подготовки естественных рамок для антропогенеза и колыбели для человека – я хочу сказать, для гарантирования существенной объективности эволюции – необходимо и достаточно одно, чтобы общий филогенез жизни (каковы бы ни были его процесс и движущая пружина) был для нас столь же отчетливо различим, как и индивидуальный ортогенез, через который на наших глазах проходит каждое живое существо, чему мы нисколько не удивляемся. Так вот, доказательство этого общего развития биосферы образуется в нашем уме почти механически и неотвратимо тем материальным рисунком, к которому мы неизбежно приходим при каждой новой попытке установить, штрих за штрихом, контуры и структуру организованного мира. Никому не придет в голову подвергнуть сомнению вихревое происхождение спиральных туманностей; или последовательное приращение частиц у кристалла или сталагмита; или наращивание пучков волокон вокруг оси стебля. Некоторые геометрические построения, на наш взгляд совершенно неподвижные, являются следом и верным признаком кинематики. Как же можно хоть на миг усомниться в эволюционном происхождении живого слоя Земли? Уступая силе анализа, жизнь расслаивается. Она до бесконечности расчленяется в анатомически и физиологически связную систему вставленных друг в друга вееров. *) Едва наметившиеся микровеера подвидов и рас. Уже более широкие веера видов и родов. Все более и более обширные веера биотов, затем пластов и далее ветвей. И, наконец, вся совокупность животного и растительного царств, образующая путем ассоциации один гигантский биот, быть может, берущий начало как один простой побег от мутовки, находящейся в глуби мегамолекулярного мира. Жизнь – простая ветвь чего-то другого... *) Разумеется, в этой игре вееров стыки можно было бы наметить иначе, чем это сделано мною, – в частности отведя больше места параллелизму и схождению. Например, четвероногие могут рассматриваться как пучок, состоящий из ряда побегов, вышедших из различных мутовок, но одинаково пришедших к формуле четвероножия. Эта полифилетическая схема, на мой взгляд, менее верно выражает факты. Но она нисколько не меняет моего основного положения, а именно, что жизнь выступает как органически сочлененный ансамбль, с очевидностью выражающий феномен роста. Сверху донизу, от самого большого до самого малого – одна и та же видимая структура, рисунок которой, подкрепленный самим распределением теней и пустот, принимает все более четкий характер и продолжается (независимо от всякой гипотезы!) в почти самопроизвольном расположении непредвиденных элементов, добавляющихся ежедневно. Каждая вновь открытая форма находит свое естественное место, фактически в данных рамках ни одна из них не является абсолютно "новой". Что еще нужно для убеждения, что все это родилось, что все это выросло?.. После этого мы можем еще годами спорить о том, как возник этот огромный организм. Убеждаясь все больше в ошеломляющей сложности его строения, мы испытываем головокружение. Как совместить этот постоянный рост с детерминизмом молекул, со слепой игрой хромосом, с кажущейся неспособностью индивидуальных приобретений передаваться по наследству? Иначе говоря, как совместить внешнюю, "финалистскую" эволюцию фенотипов с внутренней, механистической эволюцией генотипов?.. Разбирая машину на части, мы перестаем понимать, как она может двигаться вперед. Может быть и так. Но тем не менее машина перед нами, и она движется. Если химия еще не может сказать ничего вразумительного о способе образования гранитов, то можно ли сомневаться в том, что континенты беспрестанно становятся все более гранитными? Как все в универсуме, где время окончательно (я к этому еще вернусь) стало четвертым измерением, жизнь не может не быть величиной эволюционной по своей природе или размерам. Физически и исторически она соответствует некоторой функции X, определяющей положение каждого живого существа в пространстве, в длительности и по форме. Вот фундаментальный факт, который требует объяснения, но очевидность которого уже не нуждается в проверке и гарантирована от опровержения опытом в последующем. На этой ступени обобщения можно сказать, что "трансформистский вопрос" более не существует, он окончательно решен. Отныне, чтобы поколебать наше убеждение в реальности биогенеза, надо было бы подрывая всю структуру мира, вырвать с корнем древо жизни. *) *) Действительно, постольку поскольку эволюционизм выражает простую невозможность для нас воспринимать в опыте любое существо (живое или неживое) иначе, как в пространственно-временном ряду, он давно перестал быть гипотезой и стал условием (мерности), которому отныне должны отвечать в физике и биологии все гипотезы. В настоящее время биологи и палеонтологи еще спорят о конкретных путях и особенно о механизме превращений жизни: имеет ли место при возникновении новых признаков господство случая (неодарвинизм) или же факт изобретения (неоламаркизм). Но с тем общим и основным фактом, что органическая эволюция происходит как в случае любого живого существа в отдельности, так и в случае жизни в целом, ныне согласны все ученые; и это по той простой причине, что иначе они вообще не могли бы заниматься наукой... Можно лишь сожалеть (и удивляться), что. несмотря на ясность фактов. единодушие еще не доходит до признания того, что "галактика" жизненных форм вырисовывает (как это допущено нами) широкое "ортогенетическое" движение свертывания в направлении все большего возрастания сложности и сознания (см. Заключение в конце книги). |