Рефераты по вирусологии. Рефераты по вирусологии сельскохозяйственных животных Введение в вирусологию Современная таксономия вирусов
 Скачать 465.5 Kb. Скачать 465.5 Kb.
|

 Ордена Дружбы народов Российский Университет Дружбы Народов Медицинский факультет Кафедра микробиологии, вирусологии и иммунологии Рефераты по вирусологии сельскохозяйственных животных «Введение в вирусологию» «Современная таксономия вирусов» «Герпесвирусы» «Коронавирусы» Студент: А. Ю. Павлюченко Группа: СВ-32 Руководитель: Шарова И. Н. г. Москва 2004 г. Содержание Введение в вирусологию 4 Определение понятия «вирус» 4 + 4 История открытия 5 Место вирусов в биосфере 6 Борьба с вирусами – профилактика болезней 8 Современная таксономия вирусов 10 Иерархическая система классификации вирусов 10 Балтиморская классификация 12 Семейство Herpesviridae 15 История открытия 15 Структура 15 Таксономия 15 Геном 15 Репликация 16 Инфекционный ринотрахеит и пустулезный вульвовагинит КРС 17 Злокачественная катаральная горячка КРС 17 Болезнь Ауески 17 История открытия 18 Структура 18 Таксономия 19 Геном 19 Репликация 19 Литература 21 Введение в вирусологиюОпределение понятия «вирус»Несмотря на многолетнее развитие учения о вирусах, до сих пор нет общепринятого определения вирусов. Первоначально А.Львов (1957 г.) предложил определять вирусы как «строго внутриклеточные, потенциально патогенные агенты с инфекционной фазой развития,
В другом определении (С.Лурия, Дж. Дарнелл, 1967) вирусы рассматриваются как «объекты, геномы которых состоят из нуклеиновой кислоты и репродуцируются в живых клетках; используя их синтетический аппарат, они вызывают синтез специализированных структур, способных переносить геном вируса в другие клетки». В первом определении сделан акцент на внутриклеточной природе вирусов, их исключительной зависимости от метаболизма клетки-хозяина и на том, что на определенной стадии репродуктивного цикла специфический материал вируса представлен генетическим материалом - нуклеиновой кислотой. Во втором определении подчеркнуто два существенных качества вируса: 1) наличие у вируса собственного генетического материала, использующего биохимический аппарат клетки-хозяина, 2) существование у вирусов внеклеточной инфекционной фазы, представленной вирионами, которые репродуцируются- под генетическим контролем данного вируса и служат для введения генома вируса в другие клетки. Таким образом, в отличие даже от самых мелких микроорганизмов, таких как риккетсии, хламидии и микоплазмы, вирусы не имеют клеточной организации, хромосомных генов, рибосомальной системы и митохондриального аппарата (или его эквивалента для производства энергии). Таблица 1. Отличительные свойства одноклеточных микроорганизмов и вирусов
История открытияПастер был первым, кто начал (1881 г.) систематически использовать лабораторных животных в работах по изучению вируса бешенства. Его исследования положили начало введению вирусных агентов в восприимчивый организм, а также в наиболее чувствительные к заболеванию органы или ткани. В 1884 г. ученик Пастера Шамберлан изобрел керамические фильтры, позволившие освобождать от бактерий проходящие через них жидкости. В результате наблюдений Д. И. Ивановский и В. В. Половцев впервые высказали предположение, что болезнь табака, описанная в 1886 году Mayer в Голландии под название мозаичной, представляет собой не одно, а два совершенно различных заболевания одного и того же растения: одно из них - рябуха, возбудителем которого является грибок, а другое - неизвестного происхождения. Исследование мозаичной болезни табака Д.И. Ивановский продолжает в Никитинском ботаническом саду (под Ялтой) и ботанической лаборатории Академии наук и приходит к выводу, что мозаичная болезнь табака вызывается бактериями, проходящими через фильтры Шамберлана, которые, однако, не способны расти на искусственных субстратах. Возбудитель мозаичной болезни называется Ивановским то “фильтрующимися” бактериями, то просто микроорганизмами. Подчеркивая, что возбудитель мозаичной болезни табака не мог быть обнаружен в тканях больных растений с помощью микроскопа и не культивировался на искусственных питательных средах. Д. И. Ивановский писал, что его предположение о живой и организованной природе возбудителя “формировано в целую теорию особого рода инфекционных заболеваний”, представителем которых, помимо табачной мозаики, является ящур (использовав тот же метод фильтрации). Д. И. Ивановский открыл вирусы - особую форму существования жизни. Своими исследованиями он заложил основы ряда научных направлений вирусологии: изучение природы вируса, цитопаталогических вирусных инфекций, фильтрующихся форм микроорганизмов, хронического и латентного вирусоносительства. Один из выдающихся советских фитовирусологов В. Л. Рыжков писал: “Заслуги Д. И. Ивановского не только в том, что он открыл совершенно новый вид заболеваний, но и в том, что он дал методы их изучения”. 1902 г. открыт вирус гриппа кур, вызывавший массовое летальное заболевание у кур, получившее первоначально название «классическая чума птиц». Впоследствии были выделены вирусы гриппа свиней (1932 г.) и человека (1933 г.). В 1908 г. Эллерман и Банг обнаружили, что лейкоз кур передается бесклеточными фильтратами, а Швейнитц и Дорсе (1903 г.) установили вирусную природу классической чумы свиней. В 1911 г. Раус открыл вирус, вызывающий у кур злокачественные опухоли. Эти и другие наблюдения послужили основанием считать вирусы важными факторами онкогенеза. Туорт (1915 г.) и д'Эрелль (1917 г.) независимо друг от друга открыли вирусы бактерий (бактериофаги). В 1935 году У. Стенли из сока табака, пораженного мозаичной болезнью, выделил в кристаллическом виде ВТМ (вирус табачной мозаики). За это в 1946 году ему была вручена Нобелевская премия. В 1958 году Р. Франклин и К. Холм, исследуя строение ВТМ, открыли, что ВТМ является полым цилиндрическим образованием. В 1960 году Гордон и Смит установили, что некоторые растения заражаются свободной нуклеиновой кислотой ВТМ, а не целой частицей вириона. В этом же году крупный советский ученый Л. А. Зильбер сформулировал основные положения вирусогенетической теории. В 1962 году американские ученые А. Зигель, М. Цейтлин и О. И. Зегал экспериментально получили вариант ВТМ, не обладающий белковой оболочкой, выяснили, что у дефектных ВТМ частиц белки располагаются беспорядочно, и нуклеиновая кислота ведет себя, как полноценный вирус. В 1968 году Р. Шепард обнаружил ДНК-содержащий вирус. Одним из крупнейших открытий в вирусологии является открытие американских ученых Д. Балтимора и Н. Темина, которые нашли в структуре ретровируса ген, кодирующий фермент - обратную транскриптазу. Назначение этого фермента - катализировать синтез молекул ДНК на матрице молекулы РНК. За это открытие они получили Нобелевскую премию. В знак признания выдающихся заслуг Д. И. Ивановского перед вирусологической наукой Институту вирусологии АМН СССР в 1950 году было присвоено его имя, а в Академии медицинских наук учреждена премия имени Д. И. Ивановского, присуждаемая один раз в три года. Место вирусов в биосфереПо мере изучения природы вирусов в первые 50 лет после их открытия Д. И. Ивановским (1892) формировались представления о вирусах как о мельчайших организмах. Эпитет “фильтрующийся” со временем был отброшен, так как стали известны фильтрующиеся формы или стадии обычных бактерий, а затем и фильтрующиеся виды бактерий. Наиболее правдоподобной и приемлемой является гипотеза о том, что вирусы произошли из “беглой” нуклеиновой кислоты, т.е. нуклеиновой кислоты, которая приобрела способность реплицироваться независимо от той клетки, из которой она возникла, хотя при этом предусматривается, что такая ДНК реплицируется с использованием структур этой или другой клеток. На основании опытов фильтрации через градуированные линейные фильтры были определены размеры вирусов. Размер наиболее мелких из них оказался равным 20-30 нм, а наиболее крупных - 300-400 нм. Таким образом вирусы, должно быть, произошли от клеточных организмов, и их не следует рассматривать, как примитивных предшественников клеточных организмов. Положение о том, что вирусы представляют собой полноценные организмы, позволило окончательно объединить все три названных группы вирусов - вирусы животных, растений и бактерий - в одну категорию, занимающую определенное место среди живых существ, населяющих нашу планету. Тот факт, что их не удалось выращивать на искусственных питательных средах, вне клеток, не вызывал особого удивления, так как вирусы с самого начала были определены как облигатные внутриклеточные паразиты. Это свойство признавалось не уникальным, присущим только вирусам, поскольку внутриклеточные паразиты известны и среди бактерий, и среди простейших. Как и другие организмы, вирусы способны к размножению. Вирусы обладают наследственностью, воспроизводя себе подобных. Наследственные признаки вирусов можно учитывать по спектры поражаемых хозяев и симптомам вызываемых заболеваний, а также по специфичности иммунных реакций естественных хозяев или искусственных иммунизируемых экспериментальных животных. Сумма этих признаков позволяет четко определить наследственные свойства любого вируса, и даже больше - его разновидностей, имеющих четкие генетические маркеры, например: нейротропность некоторых вирусов гриппа, сниженную патогенность у вакцинных вирусов и т.п. Изменчивость является другой стороной наследственности, и в этом отношении вирусы подобны всем другим организмам, населяющим нашу планету. При этом у вирусов можно наблюдать как генетическую изменчивость, связанную с изменением наследственного вещества, так и фенотипическую изменчивость, связанную с проявлением одного и того же генотипа в разных условиях. Примером первого типа изменчивости являются мутанты одного и того же вируса, в частности термочувствительные мутанты. Примером второго типа изменчивости служит разный тип поражений, вызываемых одним и тем же вирусом у различных животных, растений и бактерий. Все вирусы по своей природе - паразиты. Они способны воспроизводить себя, но только внутри живых клеток. Обычно вирусы вызывают явные признаки заболевания. Попав внутрь клетки, они “включают” ее ДНК и, используя свою собственную ДНК или РНК, дают клетке команду синтезировать компоненты вируса. Компоненты вируса способны к спонтанному образованию вириона. В случае литического исхода вирусы “разрывают” оболочку клетки и передаются в другую клетку в виде инертных частиц. Ученные, анализируя строение вещества, до сих пор не решили: считать вирусы живыми или мертвым. Вирусы, с одной стороны, обладают способностью размножатся, наследственностью и изменчивостью, но с другой стороны, не имеют обмена веществ, и их можно рассматривать, как комплекс молекул. Вирусы как и другие организмы, характеризуются приспособляемостью к условиям внешней среды. Нужно только не забывать, что для них организм хозяина является средой обитания, поэтому многие условия внешней среды влияют на вирус опосредованно - через организм хозяина. Однако многие факторы внешней среды могут и непосредственно воздействовать на вирусы. Достаточно вспомнить уже названные термочувствительные мутанты вирусов, которые, например, размножаются при температуре 32-37 С и гибнут при температуре 38-40 С, хотя их хозяева остаются вполне жизнеспособными при этих температурных режимах. В связи с тем, что вирусы являются паразитами, они подчиняются закономерностям и к ним применимы понятия экологии паразитизма. Каждый вирус имеет круг естественных хозяев, иногда очень широкий, как, например, у мелких РНК-геномных фагов: в первом случае поражаются все млекопитающие, во втором - отдельные клоны кишечной палочки. Циркуляция вирусов может быть горизонтальной (распространение среди популяции хозяев) и вертикальной (распространение то родителей потомству). Таким образом, каждый вирус занимает определенную экологическую нишу в биосфере. Борьба с вирусами – профилактика болезнейЕще в XVIII веке Дженнер приготовил вакцину против оспы, а Пастер в XIX веке — против бешенства. Болезни, которые мы сейчас относим к вирусным, известны с глубокой древности. Оспа в прошлом была одним из наиболее распространенных и опасных заболеваний. Эпидемия, описанная в X веке до нашей эры в Китае, очень напоминала оспу. Особенно свирепой эпидемия оспы была в XVI-XVIII веках нашей эры. В отдельные годы в Европе заболевало до 12 миллионов человек, из которых более 10% погибало. В 1576 году только в Перу погибло свыше двух миллионов человек. Во время завоевания Мексики в 1520 году погибло от оспы 3,5 миллиона человек. В 1610 году в Сибири от оспы вымерла треть местного населения (В.М. Жданов и др., 1975). История борьбы с оспой вступила в новую эру, когда английский врач Эдвард Дженнер (1796 г.) проверил сделанное крестьянами наблюдение, что переболевание коровьей оспой защищает человека от заражения натуральной оспой. В 1798 году он опубликовал свой первый научный труд «Изучение причин и действия оспенной вакцины». Однако обстоятельно описанный в нем метод натолкнулся на ожесточенное сопротивление, прежде чем начать победоносное шествие по всему миру. С помощью глобальной вакцинопрофилактики к 1979 году во всем мире искоренена натуральная оспа. В начале 80-х годов XIX века Пастер открыл возможность ослабления вирулентности различных возбудителей (сибирская язва, бешенство и др.) и их использования для защиты против соответствующих болезней. В честь открытия Дженнера Пастер предложил именовать все прививки общим термином «вакцинация» (от латинского слова vacca -- корова). С тех пор под вакцинацией понимают введение различных биологических препаратов с целью создания специфического активного иммунитета. Вакцинопрофилактика справедливо считается крупнейшим достижением биологии. Характерная черта развития биологии на современном этапе - стремительное преодоление расстояния, отделяющего фундаментальные открытия от их практического применения. Этот процесс наиболее отчетливо проявляется в области разработки средств специфической профилактики вирусных заболеваний. Благодаря этому в последние годы достигнуты большие успехи в области вакцинопрофилактики многих опасных вирусных заболеваний человека и животных. Существует эффективный контроль таких грозных заболеваний, как корь, бешенство, чума свиней, чума крупного рогатого скота, чума плотоядных, ньюкаслская болезнь, болезнь Марека и многих других вирусных болезней. Наиболее ярким примером сказанному может служить борьба с полиомиелитом и ящуром. В 50-е годы нашего столетия отмечено резкое увеличение заболевания полиомиелитом в ряде стран Америки и Европы. По данным 1956 года, в США зарегистрировано более 300 тысяч инвалидов после паралитических осложнений полиомиелита. Разработка метода размножения полиовируса в культуре клеток приматов к этому времени (Эндерс и др., 1949 г.) явилась необходимым условием для создания вакцины. Сначала это была инактивированная вакцина Солка, затем — более эффективная живая вакцина Себина. В результате массовой вакцинации заболеваемость полиомиелитом в 1965 году по сравнению с 1958 годом снизилась в 50 раз. В дальнейшем заболевание встречалось в виде спорадических случаев. Начались поиски путей полного искоренения этой инфекции. Эпопея борьбы с полиомиелитом явилась разительной иллюстрацией быстрого развития и эффективного использования новых методов исследования в решении актуальных практических задач и особенно в разработке и производстве вакцин. История борьбы с полиомиелитом во многом сходна с историей борьбы с ящуром животных, протекавшей в то же самое время с широким использованием метода клеточных культур и других достижений общей вирусологии. Успешное решение этих двух глобальных проблем оказало благотворное влияние на развитие вирусологии в целом и явилось прекрасной школой для ряда поколений вирусологов. Современная таксономия вирусовВирусы в классификации группируются по таким свойствам как:
Существуют две системы классификации: Иерархическая система классификации вирусов и Балтиморская Система Классификации. Иерархическая система классификации вирусовВ 1962 Lwoff, R. W. Horne, и P. Tournier создали всестороннюю схему классификации вирусов, имеющую следующие таксоны:
Созданный позднее Международный комитет по номенклатуре вирусов использовал многие принципы данной классификации. Основа этой классификации – вирусы группируются согласно их общим свойствам, а не свойствам поражаемых ими клеток или организмов. Используются 4 главные характеристики:
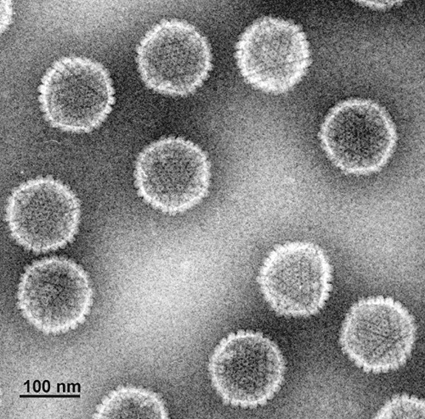
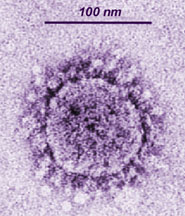
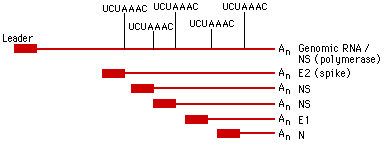
В настоящее время данная классификация актуально лишь от уровня семейства и ниже. В семейства вирусы объединены согласно эволюционному положению нуклеиновых кислот и белковых последовательностей. Все Семейства имеют суффикс -viridae, роды имеют суффикс –virus.  Рис. 1. Иерархическая система классификации вирусов Балтиморская классификацияБалтиморская система классификации вирусов основана на различиях в механизмах репликации вирусного генома, по этому принципу вирусы группируются в 7 фундаментальных групп. Любой вирус должен синтезировать положительную иРНК для синтеза собственных белков и сборки вирионов. Механизмы, предшествующие синтезу +иРНК, и отличают каждое семейство. Классификация предложена Дэвидом Балтимором и носит его имя. Класс I: Двуспиральная ДНК (папова-, адено-, герпес-, поксвирусы). 1а). Репликация происходит исключительно в ядре (папова-, адено-, герпесвирусы). Репликация этих вирусов находится в сильной зависимости от клеточных факторов. 16). Репликация происходит в цитоплазме клеток (поксвирусы). Все необходимые факторы для транскрипции и репликации кодируются вирусным геномом. Репликация относительно независима от клеточных факторов. Класс II: Односпирапьная (+) ДНК (парвовирусы). Репликация происходит в ядре. Первоначально формируются (-) цепи ДНК, которые используются как матрицы для синтеза (+) ДНК. Класс III: Двуспиральная РНК (реовирусы). Эти вирусы имеют сегментированные геномы. Каждый сегмент транскрибируется раздельно и продуцирует индивидуальные моноцистронные мРНК. Класс IV: Односпиральная (+) РНК (пикор-на-, калици-, тога-, флави-, коронавирусы). IVa). Вирусы с полицистронной мРНК (полно-, калици-, флавивирусы). Геномная РНК транслируется после инфекции, образуется полипротеин, который специфически рассекается на функциональные вирусные белки. IV6). Вирусы с комплексной трансляцией (тога-, коронавирусы). Геном транслируется в два этапа. На первом образуются ранние белки, обеспечивающие трансляцию поздних белков и последующую репликацию вирусного генома. Класс V: Односпиральные (-)РНК (ортомик-со-, парамиксо-, рабдо-, фило-, буньавирусы). Va). Сегментированные геномы (ортомиксо-, арена-, буньавирусы). На первом этапе транскрибируются геномные (-)РНК с помощью вирионной РНК-зависимой РНК-полимеразы. Образуются моноцистронные мРНК, которые служат матрицами для репликации геномной (-)РНК и трансляции вирусных белков. V6). Несегментированные геномы (парамиксовирусы). Геномные (-)РНК транскрибируются с помощью вирионной РНК-РНК-полимеразы. Образуются моноцистронные мРНК. Транслируются вирусные белки, в т.ч. вирусная транскриптаза, которая обеспечивает репликацию единой геномной (-) РНК. Класс VI: Односпиральная (+) РНК-матрица для синтеза ДНК-провируса (ретровирусы). Данный класс содержит диплоидный (+)РНК-геном, который не функционирует как мРНК, а служит матрицей для синтеза комплементарной ДНК с помощью вирионной РНК-зависимой ДНК-полимеразы. Образовавшийся ДНК-провирус интегрирует в клеточный геном и оттуда реализуется в виде моноцистронных вирусных мРНК. Класс VII. (дополнительный) (гепадновирусы). Геномная ДНК гепадновирусов (частично двуспиральная) на первом этапе достраивается до ковалентно замкнутой кольцевой молекулы. На достроенной геномной ДНК синтезируется (+)РНК, которая служит матрицей для обратной транскрипции и образования (-)-нити ДНК. После деградации (+)РНК на (-)РНК синтезируется комплементарная (+)ДНК, которая до конца не достраивается, формируя своеобразный (прерывистый) геном гепадновирусов. Для репликации ДНК-генома не требуется его интеграции в клеточную ДНК. 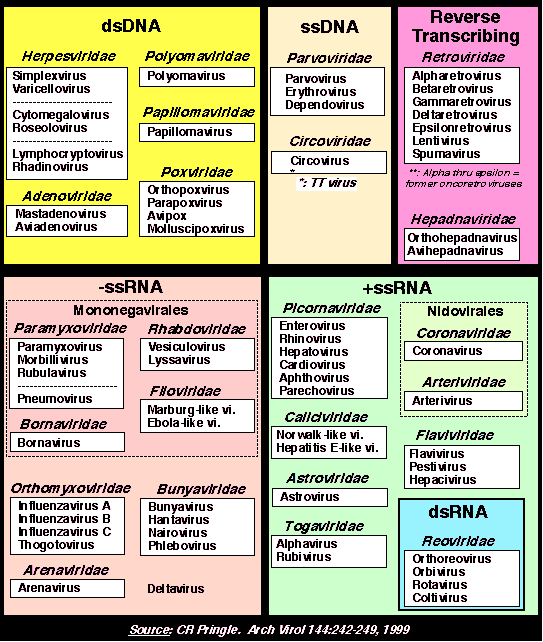 Рис. 2. Балтиморская система классификации вирусов Семейство HerpesviridaeИстория открытияНазвание произошло от греческого Herpein – «красться», т.к. инфекции хронические, латентные, рецидивирующие. Эпидемиология основных герпесвирусных инфекций ставила в тупик клиницистов много лет. В 1950 г. Burnet and Buddingh показали, что HSV может оставаться латентным после первичной инфекции до тех пор, пока не произойдет его реактивация вследствие провокации. Weller (1954) выделил VZV из везикул на коже кур. К настоящему времени известно примерно 100 герпесвирусов, из них 8 поражают человека. СтруктураРазмер: 180-200 нм Суперкапсид: Есть; содержит гликопротеиды. Тегумент: Заполненное белком пространство между суперкапсидом и капсидом Капсид: Икосаэрдический, 95-105 нм в диаметре; 162 гексагональных капсомеров. Сердцевина: Тороидальная (ДНК вокруг протеина), |