шпоры по селекции 2012 год. Селекция как наука и отрасль с х. производства
 Скачать 229.79 Kb. Скачать 229.79 Kb.
|

|
Интенсивность отбора можно определить с помощью селекционного дифференциала (S) или показателя интенсивности отбора (i). Селекционный дифференциал представляет собой разность между средней величиной признака в популяции отобранных особей (хе) и соответствующей средней его величиной в исходной популяции (хр): 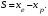 Например, если в популяции со средней высотой растений хр = 125 см отбирают все низкорослые растения со средней высотой хе = 90 см, то полученный селекционный дифференциал составит 35 см. Чем интенсивнее ведётся отбор, тем выше значение S. С помощью Sстрогость отбора по определённому признаку в различных популяциях можно сравнивать только в том случае, если изучаемые популяции имеют одинаковую изменчивость определённого признака. Этого обычно не бывает. Наследуемость. Отбор будет действенным в том случае, если хотя бы часть наблюдаемой фенотипической изменчивости признака, подлежащего отбору, обусловлена генотипически. Чем больше доля генотипической изменчивости, тем теснее связь генотипа с фенотипом и тем эффективнее отбор. Мерой доли генотипически обусловленной изменчивости в общем фенотипическом варьировании служит коэффициент наследуемости. Косвенно по коэффициенту наследуемости можно судить и о доле изменчивости, обусловленной влиянием внешней среды. Коэффициентом наследуемости в широком смысле называется отношение генотипической вариансы к общей фенотипической вариансе: 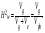 Коэффициент наследуемости может варьировать от 0 до 1. Чем выше коэффициент наследуемости, тем эффективнее отбор. Сдвиг при отборе. Коэффициент наследуемости даёт возможность предсказать результат (сдвиг) отбора. Сдвигом отбора, или респонсом R, называется наследуемая часть селекционного дифференциала S. Она равна произведению селекционного дифференциала на коэффициент наследуемости: Коэффициент наследуемости может варьировать от 0 до 1. Чем выше коэффициент наследуемости, тем эффективнее отбор. Сдвиг при отборе. Коэффициент наследуемости даёт возможность предсказать результат (сдвиг) отбора. Сдвигом отбора, или респонсом R, называется наследуемая часть селекционного дифференциала S. Она равна произведению селекционного дифференциала на коэффициент наследуемости: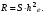 26 общие принципы отбора в селекции Отбор включает следующие этапа: 1) в исходной популяции (из исходного материала) отбирают особи по определённым критериям; 2) проводят испытания потомств отобранных растений, проверяя успех отбора. На обоих этапах соблюдают определённые требования к условиям выращивания. 1. Селекционный материал выращивают в гомогенных почвенных условиях. 2. условия выращивания должны быть репрезентативными для особенностей производственной зоны, для которой выводится сорт. 3. При выращивании исходного материала каждое растение должно иметь определённую и одинаковую для всех растений площадь питания, которая в свою очередь должна обеспечивать проведение точной оценки каждой особи исходного материала. 4. Размер площади питания должен быть близкий к принятому в производстве. 27 Виды массового отбора и его использование в селекции . Массовый отбор – отбор, при котором из исходной популяции отбирается большое число сходных по комплексу признаков лучших (элитных) растений. После лабораторной браковки урожай этих растений объединяется и высевается на следующий год на одной делянке, создавая новую популяцию. Выделяют несколько видов массового отбора: однократный; многократный; непрерывный; негативный. В селекции самоопыляющихся растений при отборе по качественным признакам в основном проводят однократный массовый отбор. В селекции перекрёстноопыляющихся растений применяют только многократный и непрерывный массовый отборы. В ряде случаев от общей массы типичных для селекционной формы или сорта растений необходимо удалить незначительное число нетипичных растений или примесь. Массовый отбор в этом случае называется негативным. Негативный массовый отбор чаще применяют на ранних этапах селекции и в семеноводстве. Процесс массового многократного отбора у перекрёстноопыляющихся культур сводится к следующему. В первый год из исходной популяции (местного сорта) отбирают лучшие элитные растения (рис. 9.3). Их урожай объединяют и на следующий год семена высевают для проведения повторного массового отбора. Часть семян урожая первого отбора высевают в сортоиспытании. Здесь результаты первого отбора сравнивают с исходной популяцией, а также с лучшим стандартным реестровым сортом. Урожай повторного массового отбора используют на посев для проведения третьего отбора и дальнейшего сортоиспытания. Если сортоиспытание дало хорошие результаты, организуют предварительное размножение нового сорта и испытание его на государственных сортоиспытательных участках. В последующие годы работу по этой схеме продолжают в таком же порядке до получения нужных результатов. Непрерывный массовый отбор используют, например, для увеличения и поддержания сахаристости у сахарной свёклы, масличности – у подсолнечника. 28 индивидуальный отбор в селекции самоопыляющих культур м-ды пересева и родословных. Индивидуальный отбор – отбор, основанный на оценке по потомству отобранных и индивидуально размножаемых лучших растений. Сущность этого метода в том, что отбирают отдельные растения и потомство каждого их них размножают в дальнейшем отдельно. Исходные родоначальные особи многократно проверяют по потомству для проверки их генотипической ценности. Потомства худших, случайно отобранных растений, выбраковывают. Путём индивидуального отбора из естественных популяций и селекционных сортов выведены, например, сорта озимой пшеницы Ульяновка и Горьковчанка; яровой пшеницы Лютесценс 62 и Минская; овса Советский и Победа; ячменя Нутанс 187 и Айхал; большинство сортов проса, гороха и чечевицы. Новый сорт овса Иртыш 13 создан путем индивидуального отбора из сорта Хармон (Канада, К-11449). Из гибридных популяций индивидуальным отбором созданы сорта озимой пшеницы Безостая 1 и Кавказ, яровая пшеница Саратовская 29 и многие другие современные сорта (Терция, Соната, Дуэт, Эритроспермум 59, Нива 2). Индивидуальный отбор у самоопылителей чаще бывает однократным и сводится к последовательной оценке потомств однажды отобранных элитных растений во всех звеньях селекционного процесса. Например, индивидуальным отбором в F2 создан сорт яровой мягкой пшеницы Омская 18, в F3 – сорта ячменя Омский 91 и Омский голозерный 1, сорт гороха Батрак. Сорт яровой мягкой пшеницы Чернява 13 создан путем отбора из гибридной популяции F5. Процесс однократного индивидуального отбора у самоопылителей сводится к следующему. В первый год из исходной популяции отбирают и отдельно обмолачивают лучшие растенияСемена всех оставшихся после браковки элитных растений на следующий год высевают отдельно в селекционном питомнике. Здесь бракуют худшие линии и оставляют лучшие. Каждая линия, высеваемая в селекционном питомнике (контрольном питомнике), получает определённый номер. Под этим номером она проходит все дальнейшие испытания и размножения 29 особенности отбора в селекции перекрестноопыляющих культур, м-д половинок, индивидуально семейного и семейно-группового отбора. У перекрёстноопыляющихся видов индивидуальный отбор используют для улучшения как полигенных признаков со средней наследуемостью, так и моногенных доминантных признаков. Так как элитные растения аллогамных видов, как правило, гетерозиготны, в их потомствах проявляется генотипическая изменчивость и дальнейший отбор может оказаться эффективным. К основным проблемам, ограничивающим применение индивидуального отбора у перекрёстников, относятся: 1) неконтролируемое переопыление растений между собой; 2) проявление инцухт-депрессии у потомства при близкородственном опылении. Метод половинок – приём использования многократного индивидуального отбора в селекции перекрёстноопыляющихся культур, при котором урожай каждого элитного растения делится на две части (половинки) Одна часть семян высевается в селекционном питомнике, а другая – сохраняется в резерве. На следующий год в целях исключения нежелательного влияния отцовских родительских форм селекционный питомник засевается с применением изоляции семенами резервных половинок тех растений, которые положительно проявили себя в предыдущем году. В питомнике предварительного размножения переопыление допускается только между теми линиями, показатели которых были выше для всех испытанных линий. Путём применения этого метода устраняется нежелательное влияние отцовских родительских форм.Индивидуально-семейный отбор – вид индивидуального отбора у перекрёстноопылителей, при котором семена каждого элитного растения высеваются по семьям на отдельных изолированных площадках Переопыление растений происходит всё время только в пределах семьи. При индивидуально-семейном отборе быстро достигается выравнивание, усиление и закрепление нужных признаков, но в силу близкородственного оплодотворения наблюдается в той или иной степени выраженная инбредная депрессия. Методом индивидуально-семейного отбора созданы многие сорта ржи и гречихи. Например, петем индивидуально-семейного отбора из гибридной популяции Харьковская 88 х Чулпан создан диплоидный сорт озимой ржи Ирина (2005). Сорт озимой тетраплоидной ржи Сибирь (1999) Метод семейно-группового отбора сочетает преимущества индивидуального и массового отборов. Семейно-групповой отбор – вид индивидуального отбора у перекрёстноопылителей, при котором родоначальные элитные растения разбиваются на несколько групп по относительно сходным хозяйственно-биологическим признакам. Каждая группа семей высевается на изолированной площадке. Переопыление потомств растений происходит в пределах группы. При использовании семейно-группового отбора выравненность потомств достигается медленно, но устраняется негативный эффект близкородственного разведения. Примером использования данного метода в селекции является сорт диплоидной гречихи Наташа (2004), который был выведен многократным семейно-групповым отбором из гибридной популяции, полученной при свободном переопылении сорта Ирменка и скороспелых крупноплодных форм. 30 особенности отбора в селекции вегетативно –размнож культур. При работе с вегетативно размножающимися культурами (картофель, банан, многие цветочные, все плодовые и ягодные культуры, лесные породы) любое отобранное ценное родоначальное растение независимо от того, гомозиготное оно или гетерозиготное, может быть широко размножено вегетативным путём и выпущено в качестве нового сорта. Это позволяет на любом этапе работы выделить ценные в селекционном отношении элитные растения и путём вегетативного размножения закрепить их свойства1. В естественной популяции вегетативно размножающегося вида встречается ясно выраженная генетическая изменчивость. В этом случае отбирают элитные растения и вегетативно размножают каждое из них, а затем последовательно испытывают их потомства, т.е. клоны. Испытание клонов проводят так же, как и испытание генеративных потомств, только вместо генеративного размножения происходит вегетативное. 31 понятие о мутациях , классификация мутаций .з-чи и преимущества мутациооной селекции. Генетические структуры клетки способны изменяться под влиянием различных физических и химических факторов. Генетически стойкие изменения в генах и хромосомах называются мутациями. Новый организм с изменённым признаком вследствие мутирования гена или перестройки хромосомы называют мутантом. Мутации могут быть полезными, вредными или нейтральными для организма. Большинство из них вредные или даже летальные, например хлорофильные мутации. Мутагенами называют различные физические и химические факторы, с помощью которых получают индуцированные мутации. Из них используют главным образом различного типа излучения и химические вещества. В селекции с помощью индуцированного мутагенеза можно решать следующие задачи: 1) обеспечение изменчивости с широким спектром мутаций и высокой частотой их проявления в целях получения исходного материала для отбора;2) получение мутантов со специфическими изменениями отдельных признаков в целях исправления отдельных дефектов сортов. При этом другие хозяйственно важные признаки должны остаться без изменений;3) увеличение рекомбинации генов и разрыв нежелательного сцепления генов; 4) осуществление переноса фрагментов хромосом одного вида в хромосомы других видов при отдалённой гибридизации; 5) получение гомозиготных мутантов путём воздействия на гаплоиды излучениями и последующего удвоения у них числа хромосом и т. д. Преимущества метода мутагенеза перед другими методами селекции.1. при улучшении одного или многих признаков существенно не перестраивается генотип улучшаемого сорта. При мутагенезе возникает меньше гетерозиготных локусов в первом поколении, поэтому за более короткий срок можно получить константно не расщепляющиеся линии. 2. В результате мутаций у растений могут проявляться новые, ранее неизвестные, полезные признаки, для передачи которых гибридизация является неэффективной. Общий механизм, добавляющий большое число совершенно новых полезных признаков, заключается в том, что мутагены индуцируют перемены таких генов, которые появляются в спонтанном порядке крайне редко или не отмечаются вообще. Если мутанты имеют отдельные положительные отклонения, их используют в скрещиваниях с лучшими сортами или другими мутантами.3. Действие мутагенов на генетический аппарат более упорядочено, чем при спонтанном мутагенезе. Часть мутагенов совсем не вызывает хромосомных перестроек.4. Мутагенезом можно вызывать взаимные транслокации хромосом у отдалённых гибридов F1, когда естественная рекомбинация не даёт желаемого эффекта. При внутривидовой гибридизации частота рекомбинации тесно сцепленных генов может быть значительно повышена мутагенными факторами. Этим же устраняются нежелательные корреляции между признаками: например, между урожайностью и склонностью к полеганию, поскольку хромосомные перестройки нарушают сцепление генов, определяющих коррелирующие признаки. 32 виды и источники излучений, используемые в селекции . Излучения, вызывающие мутации, бывают двух видов: ионизирующие и неионизирующие.Среди ионизирующих излучений в селекционной работе наиболее широко применяют: 1) рентгеновское излучение; 2) гамма-излучение; 3) нейтронное излучение.Ионизирующими они называются потому, что способны превращать атомы и молекулы в электрически заряженные частицы – ионы.Рентгеновское излучение стали использовать в селекции для получения мутаций раньше всех других источников. Рентгеновские аппараты имеются во многих учреждениях, легко управляются, на них удобно работать с любыми органами растения, несложно изменять дозы излучения. Частота появления мутаций зависит от дозы излучения. Существует положительная линейная зависимость между дозой излучения и частотой мутаций. Однако эта зависимость наблюдается в основном в интервале 100%-ной выживаемости. Установлено, что сорта разной плоидности обладают неодинаковой радиочувствительностью и разным спектром мутаций. Например, у гречихи тетраплоиды значительно более устойчивы к гамма-излучению, чем диплоиды. Было показано, что с увеличением уровня плоидности у пшеницы частота хлорофильных мутантов уменьшается, а морфологических и других видимых мутантов возрастает. Кроме этого, различные сорта одного вида с одинаковым набором хромосом могут иметь разную радиочувствительность. Например, у сортов гороха Торсдаг, Рамонский 77, сои Кубанской 276, яровой пшеницы Теремок воздействие гамма-лучей на семена обусловило появление значительного количества мутаций с широким спектром, в том числе по скороспелости, продуктивности, склонности к полеганию. В тех же опытах абиссинская форма гороха, карликовый сорт сои, сорт ярового ячменя Винер оказались маломутабильными. Поскольку разные сорта часто отличаются по чувствительности к мутагенам и имеют неодинаковый спектр мутаций, для получения желаемых результатов нужно вовлекать в работу больше сортов.При высоких дозах излучения, дающих сильный повреждающий эффект, доля хозяйственно-ценных мутаций меньше, чем при средних дозах. Поэтому в селекционных целях рекомендуется использовать дозы излучения в 1,5–2 раза ниже критических Генетически эффективным неионизирующим излучением считают ультрафиолетовое (УФ) излучение. Оно имеет значительно большую длину волны (200–400 нм), чем ионизирующие излучение, и меньшую энергию. При его использовании происходит только возбуждение молекул. Проникающая способность УФ-излучения очень мала. Поэтому его можно использовать только для обработки пыльцевых зёрен. УФ-излучение вызывает высокую частоту мутаций, особенно в пределах длины волны, которая поглощается ДНК (260–265 нм). Например, при обработке пыльцы кукурузы УФ-лучами с длиной волны 253 нм в дозе 104 Дж/м2 наблюдается угнетение оплодотворения, а при увеличении дозы до 105–106 Дж/м2 пыльца становится полностью стерильной. 33 хим мутагены, конц и экспозиции, способы обработки . Химическими мутагенами можно обрабатывать сухие и проросшие семена, черенки, клубни, луковицы, инъекцировать эти вещества в стебель растений перед вступлением их в генеративную фазу и т. д. Продолжительность обработки семян варьирует от 3 до 18 ч. Концентрация мутагенов для каждой культуры и сорта устанавливается в специальных предварительных опытах. Ориентировочно рекомендуют следующие концентрации мутагенов для обработки сухих семян: этиленимин – 0,01–0,06 (% по объёму); ЭМС – 0,1–0,5; гидроксиламин – 1,5–3,0 и т. д. Установлено, что с увеличением концентрации мутагена до определённого уровня частота жизнеспособных мутаций возрастает, затем происходит её снижение в результате гибели клеток, в которых возникли изменения при повышении концентрации мутагена сверх оптимальной величины. Следовательно, в селекционной работе использование высоких концентраций мутагенов нецелесообразно. Они не должны быть и слишком низкими. Во многих случаях химические мутагены оказываются эффективнее физических. 1. Обработка семян парами химических мутагенов («обработка в газовой фазе»). Метод разработан в отделе химического мутагенеза под руководством И.А. Рапопорта. Он заключается в том, что сухие или проросшие семена помещают в эксикатор, куда добавляют несколько капель хорошо испаряющегося химического мутагена. Семена поглощают пары мутагена и подвергаются воздействию. Метод оказался эффективным для 1,4-бисдиазоацетилбутана, этиленимина и других быстро испаряющихся мутагенов. |