физиология. физиология 20-30. Суммация означает сложение отдельных одиночных сокращений, ведущее к увеличению интенсивности общего сокращения мышцы. Суммация осуществляется двумя путями Путем увеличения числа моторных единиц, сокращающихся одновременно, что называют суммацией сокращения многих волокон
 Скачать 0.96 Mb. Скачать 0.96 Mb.
|

|
Восстановительные процессы структуры мембраны и функции нервно-мышечного синапса после передачи возбуждения После завершения синаптической передачи в нервно-мышечном синапсе ионы Са2+ с помощью Са2+-АТФ-азы первично активным транспортом выводятся из цитоплазмы нервного окончания во внеклеточную среду. Продукты расщепления ацетилхолина захватываются белками-транспортерами пресинаптической мембраны и служат далее для восстановления ацетилхолина в терминали. В нервном окончании образуются новые везикулы и заполняются вновь синтезированными молекулами ацетилхолина. В постсинаптической клетке, благодаря работе Na+/К+-АТФазы, восстанавливаются ионные градиенты Na+ и К+, в результате возбудимость постсинаптической мембраны возвращается к исходному уровню. Инактивация медиатора. После того как медиатор подействовал на рецепторы, он должен быть удален из синаптической щели. Существует 3 способа удаления, или инактивации медиатора: 1. Диффузия из синаптической щели в окружающие ткани; 2. Обратный захват пресинаптическим окончанием целых молекул медиатора или ее фрагментов; 3. Ферментативный распад. Для нервно-мышечного синапса самый эффективный способ инактивации медиатора – это ферментативный распад: - требует фермента ацетилхолинэстеразы; - самый быстрый способ, обеспечивает передачу импульсов достаточно высокой частоты. 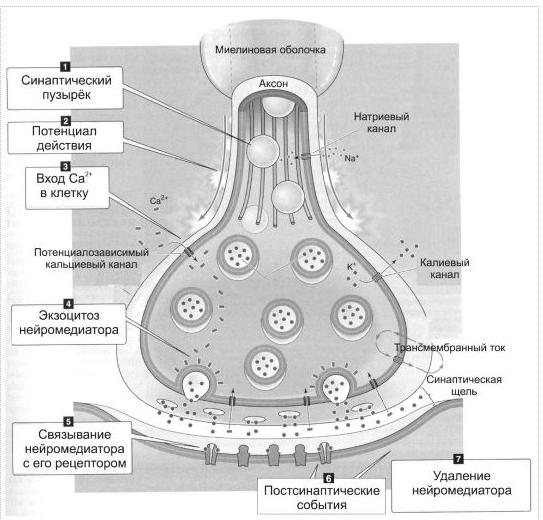 Физиологические свойства нервно-мышечного синапса. 1. Одностороннее проведение возбуждения. 2. Синаптическая задержка. 3. Низкая лабильность. 4. Высокая утомляемость. 5. Высокая чувствительность к некоторым ядам и фармакологическим препаратам. Одностороннее проведение возбуждения обусловлено направлением потока медиатора – он выделяется из пресинаптического окончания и действует на постсинаптические рецепторы. Синаптическая задержка обусловлена сравнительно длительным временем, необходимым для выделения медиатора, его диффузии к рецепторам, активации рецепторов и последующих постсинаптических процессов. Низкая лабильность. В нервных волокнах лабильность ограничена коротким временем периода рефрактерности (около 1 мс), лабильность нервных волокон высока. Синапсы же становятся готовы к проведению очередного сигнала лишь после того, как будет инактивирована очередная порция медиатора, на что требуется достаточно большое время, т.е. лабильность синапса ограничена. Следовательно, синапсы могут проводить лишь импульсы низкой частоты, т.е. обладают низкой лабильностью. Высокая утомляемость заключается в снижении величины реакции клетки при длительном раздражении. В синапсе при длительном раздражении могут истощаться запасы медиатора, и тогда развивается утомление (нерв практически неутомляем). Высокая химическая чувствительность синапса обусловлена участием в передаче сигнала химических соединений и реакций (фосфорорганические вещества являются ингибиторами холинэстеразы, кураре - препятствует развитию потенциала концевой пластинки; ботулинус - блокирует высвобождение медиатора; змеиный яд -блокирует субсинаптические рецепторы). Фармакологическая коррекция работы мионеврального синапса. Особенности синаптической передачи возбуждения определяют возможные пути ее коррекции. 1. Воздействие на скорость синтеза медиатора: а) через изменение активности ферментов (например, ацетилхолин-трансферазы); б) через изменение (угнетение) захвата холина. 2. Воздействие на секрецию медиатора и выход в синаптическую щель путем: а) угнетения проведения потенциала действия по пресинаптической области новокаином (при этом увеличивается выход калия, что приводит к гиперполяризации и снижению возбудимости); б) удаления кальция из среды; в) добавления магния или ботулинического токсина (в эксперименте). Например, белки экзоцитоза - синтаксин, SNAP-25 и синаптобревин — мишени ботулинического токсина, необратимо подавляющего слияние синаптических пузырьков с пресинаптической мембраной. 3. Воздействие на рецепторы постсинаптической мембраны: блокирование Н-холинорецепторов: - нервно-мышечных синапсов с помощью кураре и курареподобных веществ (д-тубокурарин). Например, кураре и курареподобные вещества (диплацин, тубокурарин) обратимо связываются с N-холинорецепторами постсинаптической мембраны, блокируют действие на нее ацетилхолина и передачу в синапсе. 4. Воздействие на ингибиторы медиаторов (увеличение или уменьшение активности ацетилхолинэстеразы). Например, уменьшение активности ацетилхолинэстеразы приводит к стойкой деполяризации постсинаптической мембраны, в результате чего блокируется передача возбуждения через синапс, что может привести к гибели организма. Этот механизм лежит в основе действия фосфорорганических соединений, применяемых в качестве отравляющих веществ или инсектицидов (дихлофос, хлорофос и др.). Некоторые фармакологические препараты, например прозерин, подавляют активность ацетилхолинэстеразы, способствуя умеренному накоплению ацетилхолина и облегчению синаптической передачи, что используется в лечебной практике. 24. Структурно-функциональные особенности скелетных мышц. Понятие о моторной единице. Виды моторных единиц. С функциональной точки зрения мышца состоит из двигательных единиц. Двигательная единица (ДЕ) — это понятие структурно-функциональное. В состав отдельной ДЕ входит мотонейрон и иннервируемый его аксоном комплекс мышечных волокон. Мышечные волокна, объединенные в одну ДЕ, разбросаны среди других мышечных волокон, принадлежащих другим ДЕ, и изолированы от последних. Отдельные мышцы включают в себя разное количество ДЕ. 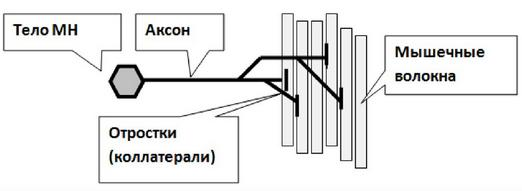 В зависимости от морфологических особенностей мотонейрона и мышечных волокон, ДЕ подразделяются на - малые, - средние; - большие. |