ИИП по биологии 11 класс. ИИП Свиридов Н. Создание пособия по решению генетических задач. Свердловской области итоговый индивидуальный проект Создание пособия по решению генетических задач
 Скачать 0.65 Mb. Скачать 0.65 Mb.
|

|
Полное доминирование Полное доминирование (рис.3.4.) — это вид взаимодействия аллельных генов, при котором фенотип гетерозигот не отличается от фенотипа гомозигот по доминанте, то есть в фенотипе гетерозигот присутствует продукт доминантного гена. 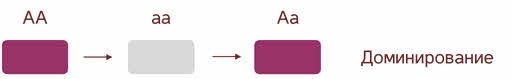 Рис.3.4. Полное доминирование Полное доминирование широко распространено в природе, имеет место при наследовании, например, окраски и формы семян гороха, цвета глаз у человека, резус-фактора и многое другое. Наличие резус-фактора эритроцитов обусловливается доминантным геном Rh (эр аш). То есть генотип резус-положительного человека может быть двух видов: или Эр-большое аш-малое Эр-большое аш-малое (RhRh), или Эр-большое аш-малое эр-малое аш-малое (Rhrh); а генотип резус-отрицательного человека только эр-малое аш-малое эр-малое аш-малое (rhrh). На рис.3.5. изображена схема наследования резус-фактора крови человека. 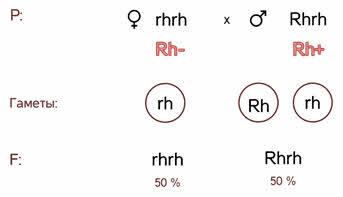 Рис.3.5. Наследование резус-фактора эритроцитов Если, например, мать — резус-отрицательная, а отец резус-положительный и гетерозиготен по этому признаку, то при данном типе брака с одинаковой вероятностью может родиться как резус-положительный, так и резус-отрицательный ребёнок. Неполное доминирование Это вид взаимодействия аллельных генов, при котором фенотип гетерозигот отличается как от фенотипа гомозигот по доминанте, так и от фенотипа гомозигот по рецессиву и имеет среднее (промежуточное) значение между ними. На рис.3.6., 3.7. показана схема неполного доминирования признака. 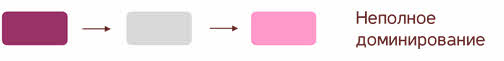 Рис.3.6. Так как у одного родительского растения оба аллеля (AA) кодируют ферменты, ответственные за синтез красного пигмента. А у второго с белыми цветками аллели (аа) содержат мутацию в результате которой фермент теряет активность, и окраска цветков получается белой. 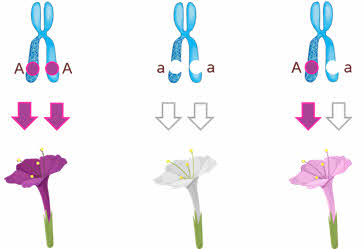 Рис.3.7. Поэтому у растений с розовыми цветками с генотипом Aа только один из аллелей кодирует фермент, который ответственный за красный цвет. Кодоминирование Это вид взаимодействия аллельных генов, при котором фенотип гетерозигот отличается как от фенотипа гомозигот по доминанте, так и от фенотипа гомозигот по рецессиву, и в фенотипе гетерозигот присутствуют продукты обоих генов.(рис.3.8.) Рис.3.8. Кодоминирование наблюдается при формировании, например, IV группы крови системы (а бэ ноль) (АВ0) у человека. Для того чтобы представить, как происходит наследование групп крови у человека, можно посмотреть рождение детей с какой группой крови возможно у родителей, имеющих один — вторую, другой — третью группы крови и являющихся гетерозиготными по этому признаку. Аллельное исключение  Аллельное исключение наблюдается при отсутствии одного из пары генов. В этом случае в фенотипе присутствует продукт другого гена. Например, при гемизиготности, делеции, гетерохроматизации участка хромосомы, в котором находится нужный ген. Перейдём к изучению взаимодействия неаллельных генов. Комплементарность или дополнительное взаимодействие Это такой тип взаимодействия неаллельних генов, когда один доминантный ген дополняет действие другого неаллельного доминантного гена, и они вместе определяют новый признак, который отсутствует у родителей. Причём соответственный признак развивается только в присутствии обоих неаллельних генов.  Например, при скрещивании двух чистых линий душистого горошка, которые имеют белые цветки у гибридов первого поколения все цветки будут иметь пурпурную окраску. Почему так происходит? Дело в том, что в данном случае доминантные неаллельные гены А-большое и В-большое, каждые в отдельности кодируют недостаточное количество красного пигмента антоциана для окраски цветка. И только при наличии обоих неаллельных доминантных генов А-большое и Бэ-большое, которые кодируют достаточное количество пигмента цветки окрашиваются в красный цвет. По принципу комплементарности наследуется окраска плодов у перца. Цвет плодов обеспечивается двумя типами пигментов – хлорофиллом и пигментами красного и жёлтого цвета. В зависимости от их сочетания плоды могут иметь четыре различных варианта окраски: красные, коричневые (бурые), жёлтые и зелёные. Доминантный аллель А-большое контролирует образование красного пигмента, тогда как рецессивный аллель а-малое жёлтого пигмента. Доминантный аллель Бэ-большое блокирует синтез хлорофилла, не влияя на образование пигментов, тогда как рецессивный аллель бэ-малое не влияет на его синтез и такие растения синтезируют хлорофилл нормально. При скрещивании растений с коричневыми плодами и растений с жёлтыми плодами в первом поколении все потомство будет иметь красные плоды. А при скрещивании гетерозиготных растений перца из первого поколения во втором поколении будет наблюдаться расщепление 9 к 3 к 3 к 1. Эпистаз Это вид взаимодействия неаллельных генов, при котором одна пара генов подавляет (не даёт проявиться в фенотипе) другую пару генов.(рис.3.9)  Рис.3.9. Угнетение могут вызывать как доминантные, так и рецессивные гены, и в зависимости от этого различают эпистаз доминантный и рецессивный. Ген-подавитель называют эпистатичным, подавляемый ген — гипостатичным. Доминантный эпистаз характерен для окраски плодов у тыквы. У этого растения известно три цвета плодов: белая, жёлтая и зелёная. Доминантный аллель гена А определяет жёлтую окраску тыквы, рецессивный аллель а-малое – зелёный цвет. Второй ген Бэ-большое проявляет эпистатическое действие – подавляет образование пигмента как жёлтого, так и зелёного цвета, делая их белыми. Рецессивный аллель бэ-малое – не влияет на проявление окраски плодов тыквы. При скрещивании растений с белыми и зелёными плодами, все потомство первого поколения будет белым. А во втором поколении расщепление признаков будет соответствовать формуле 12 к 3 к 1. В этом случае рецессивная гомозигота (a-малое a-малое бэ-малое бэ-малое) фенотипически отличима от одного из гетерозиготных классов. Таким образом растения, имеющие в своём генотипе хотя бы один доминантный ген Бэ-большое, будут образовывать белые плоды независимо от того, какие аллели гена А-большое будут присутствовать в их генотипах. Полимерное_действие_генов'>Полимерное действие генов Часто признаки в организмах могут быть выражены слабее или сильнее – например вес, удойность, плодовитость. 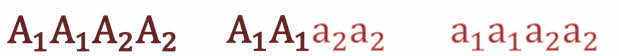 Эти признаки определяются несколькими генами, действие которых суммируется. Полимерия ─ это вид взаимодействия двух и более пар неаллельных генов, доминантные аллели которых однозначно влияют на развитие одного и того же признака. Полимерные гены обозначаются одной буквой, аллели одного локуса имеют одинаковый цифровой индекс. Полимерное действие генов может быть кумулятивным и некумулятивным.  Рис.3.10. При кумулятивной полимерии интенсивность значения признака зависит от суммирующего действия генов: чем больше доминантных аллелей, тем больше степень выраженности признака. При некумулятивной полимерии количество доминантных аллелей на степень выраженности признака не влияет, и признак проявляется при наличии хотя бы одного из доминантных аллелей. Приведём пример кумулятивной полимерии (рис.3.10.) Бурый цвет зерна пшеницы обусловлен доминантными генами из двух пар аллелей – А1 и А2. Поэтому самые яркие семена имеют растения, у которых четыре доминантных гена А1 А1 А2 А2. У растений с генотипом А1 а1 а2 а 2 или а 1 а 1А 2 а2 – зерно окрашено слабо. У растений с генотипом а 1 а 1 а 2 а 2 − зерно не окрашено. Например, при скрещивании растений с ярко бурыми семенами и растения с неокрашенными семенами в первом поколении все организмы получаются со слабо окрашенными семенами, так как им досталась половина доминантных генов от первого родителя. При скрещивании гетерозиготных растений из первого поколения происходит вот такое расщепление.  Помимо двух групп взаимодействия генов: между аллельными генами и между неаллельными генами. Существует множественное действие одного гена − плейотропность. Плейотропия — это множественное действие генов. То есть когда 1 ген влияет на несколько признаков. Например, у дрозофилы ген белого цвета глаз одновременно влияет на цвет тела, длины крыльев и снижение плодовитости. Плейотропное действие генов впервые было обнаружено Грегором Менделем, который установил, что у растений с пурпурными цветками всегда имелись красные пятна в пазухах листьев, а семенная кожура была серого цвета. То есть развитие этих признаков определяется действием одного наследственного фактора (гена). У человека встречается рецессивная наследственная болезнь — серповидно-клеточная анемия. Первичным дефектом этой болезни является замена одной из аминокислот в молекуле гемоглобина, что приводит к изменению формы эритроцитов (рис.3.11). 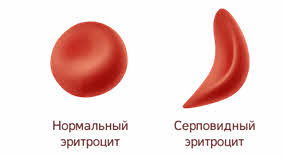 Рис.3.11.Строение эритроцита при серповидно- клеточной анемии. Одновременно с этим возникают нарушения в сердечно-сосудистой, нервной, пищеварительной и выделительной системах. Это приводит к тому, что гомозиготный по этому заболеванию ребёнок погибает в детстве. Плейотропия широко распространена. Изучение действия генов показало, что плейотропным эффектом, очевидно, обладают многие, если не все, гены. Таким образом, выражение «ген определяет развитие признака» в значительной степени условно, так как действие гена зависит от других генов — от генотипической среды. На проявление действия генов влияют и условия окружающей внешней среды. Следовательно, генотип является системой взаимодействующих генов. Глава 4. Сцепление генов Сцепление генов – это совместное наследование генов, расположенных в одной и той же хромосоме. Количество групп сцепления соответствует гаплоидному числу хромосом, то есть у дрозофилы 4. Природу сцепленного наследования объяснил Морган с сотрудниками. В качестве объекта исследования они избрали плодовую муху дрозофилу, которая оказалась очень удобной моделью для изучения данного феномена, так в клетках её тела находится только 4 пары хромосом и имеет место высокая скорость размножения (в течение года можно исследовать более 20-ти поколений). Итак, сцепленными признаками называются признаки, которые контролируются генами, расположенными в одной хромосоме. Естественно, что они передаются вместе в случаях полного сцепления (закон Моргана). Полное сцепление встречается редко, обычно – неполное, из-за влияния кроссинговера (перекрещивания и обмена участками гомологичных хромосом в процессе мейоза). То есть, гены одной хромосомы переходят в другую, гомологичную ей. Частота кроссинговера зависит от расстояния между генами. Чем ближе друг к другу расположены гены в хромосоме, тем сильнее между ними сцепление и тем реже происходит их расхождение при кроссинговере, и, наоборот, чем дальше друг от друга отстоят гены, тем слабее сцепление между ними и тем чаще возможно его нарушение. 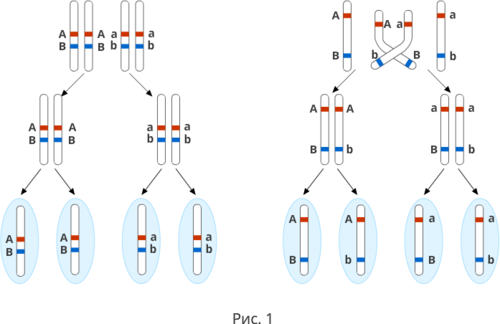 Рис.4.1 На рис.4.1. слева: расстояние между генами А и В маленькое, вероятность разрыва хроматиды именно между А и В невелика, поэтому сцепление полное, хромосомы в гаметах идентичны родительским (два типа), других вариантов не появляется. На рис.4.1. справа: расстояние между генами А и В большое, повышается вероятность разрыва хроматиды и последующего воссоединения крест-накрест именно между А и В, поэтому сцепление неполное, хромосомы в гаметах образуются четырёх типов – 2 идентичные родительским (некроссоверные) + 2 кроссоверных варианта. Количество разных типов гамет будет зависеть от частоты кроссинговера или расстояния между анализируемыми генами. Расстояние между генами исчисляется в морганидах – единицах расстояния между генами, находящимися в одной хромосоме. 1 морганида соответствует 1 % кроссинговера. Такая зависимость между расстояниями и частотой кроссинговера прослеживается только до 50 морганид. Частота кроссинговера между определенной парой генов – довольно постоянная величина (хотя радиация, химические вещества, гормоны, лекарства влияют на нее; например, высокая температура стимулирует кроссинговер). Пример, основанный на опытах Моргана  Рис. 4.2. Гены, расположенные в одной хромосоме, наследуются совместно. Фенотипы А – серое тело, нормальные крылья (повторяет материнскую форму). Б – тёмное тело, короткие крылья (повторяет отцовскую форму). В – серое тело, короткие крылья (отличается от родителей). Г – тёмное тело, нормальные крылья (отличается от родителей). В и Г получены в результате кроссинговера в мейозе. Если скрестить мушку дрозофилу, имеющую серое тело и нормальные крылья (на рисунке самка), с мушкой, обладающей тёмной окраской и зачаточными (короткими) крыльями (на рисунке самец), то в первом поколении гибридов все мухи будут серыми с нормальными крыльями (А). Это гетерозиготы по двум парам аллельных генов, причём ген, определяющий серую окраску брюшка, доминирует над тёмной окраской, а ген, обусловливающий развитие нормальных крыльев, - доминирует над геном недоразвитых крыльев (рис.4.2.). При анализирующем скрещивании гибрида F1 с гомозиготной рецессивной дрозофилой (Б) подавляющее большинство потомков F2 будет сходно с родительскими формами. Это происходит потому, что гены, отвечающие за серое тело и нормальные крылья– сцепленные гены, также как и гены, отвечающие за тёмное тело и короткие крылья, т. е. они находятся в одной хромосоме. наследование сцепленных генов называют сцепленным наследованием. Сцепление может нарушаться. Это доказывает наличие особей В и Г на рисунке 4.2., т. е. если бы сцепление не нарушалось, то этих особей бы не существовало, однако они есть. Это происходит в результате кроссинговера, который и нарушает сцепленность этих генов. 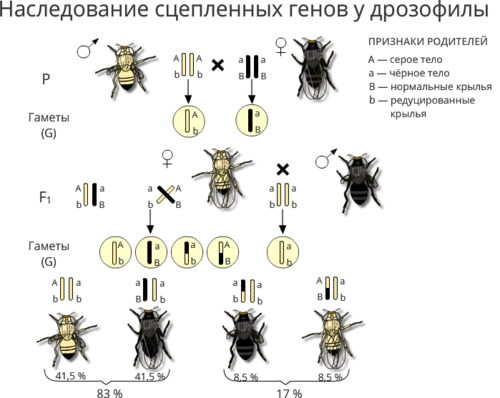 Рис. 4.3. На рисунке4.3. опыт Моргана отображен подробно. Несцепленное наследование: два гена находятся в разных хромосомах, гетерозигота с равной вероятностью дает четыре типа гамет: 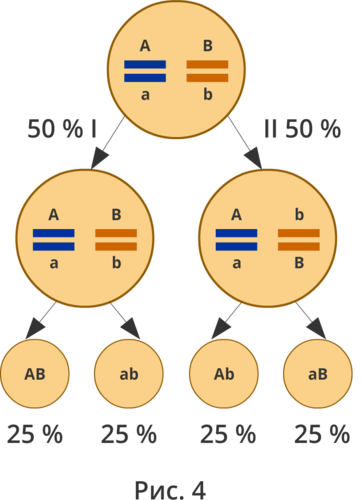 Сцепленное наследование: два гена находятся в одной хромосоме. а) При полном сцеплении гетерозигота дает только два типа гамет. б) При неполном сцеплении гетрозигота дает четыре типа гамет, но не с равной вероятностью. . 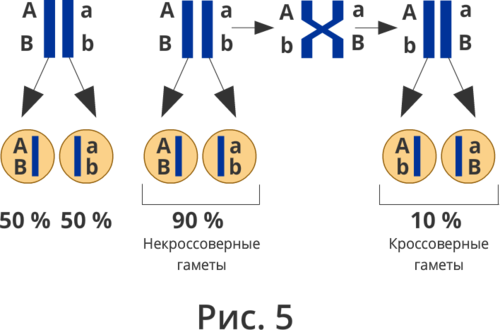 Рис.4.5. На вышесказанном строится хромосомная теория наследственности Моргана: 1. Гены находятся в хромосомах и расположены в линейной последовательности на определенных расстояниях друг от друга. 2. Гены, расположенные в одной хромосоме, составляют группу сцепления. Число групп сцепления равно гаплоидному числу хромосом. Признаки, гены которых находятся в одной хромосоме, наследуются сцепленно (т. е. в тех же сочетаниях, в которых они были в хромосомах исходных родительских форм). 3. Новые сочетания генов, расположенных в одной паре хромосом, могут возникать в результате кроссинговера в процессе мейоза. Частота кроссинговера зависит от расстояния между генами. 4. Учитывая линейное расположение генов в хромосоме и частоту кроссинговера как показателя расстояния между генами, можно построить карты хромосом. За единицу расстояния между генами принята частота кроссинговера равная 1 % (морганида, сантиморган, сМ). Заключение В ходе работы над проектом я больше узнал о жизни учёного – Грегора Менделя, этот выдающийся учёный положил начало одной из самых важных наук в биологии в наши дни. Он был не признан при жизни, но это не мешало ему продолжать проводить опыты, а нам по праву считать его отцом генетики. Я ещё раз изучил всё, что было представлено в курсе 9-11 классов, таким образом, углубив и освежив свои знания. Задачи, подобранные в пособии по разным уровням сложности, позволяют практиковаться, как тем, кто собирается сдавать экзамен по биологии, а возможно, и связать свою дальнейшую жизнь с этим увлекательным предметом, так и простым ученикам, проходящим данную тему исключительно на уроках в ознакомительных целях. Эти знания позволили мне более глубоко познакомиться с данным материалом, и применять его в нестандартных ситуациях. Поиск новой информации из различных печатных источников, а также в сети Интернет расширил мои знания по биологии. Во время составления продукта, я также расширил свои знания, узнав виды методических пособий и их назначение. Выбрав наиболее подходящий под заданную тему вид, отобрал задачи, решить которые можно несколькими способами. Разработал макет и составил методическое пособие по теме: «Решение генетических задач». Таким образом, я получил удобное и наглядное методическое пособие, которым смогу пользоваться, как я, так и другие школьники при подготовке к контрольным или экзаменам, или же просто, чтобы освежить в памяти данную тему. |