Гистология реферат. Тема 1 клетка Вопрос Химический состав, организация плазмолеммы. Функции плазмолеммы
 Скачать 3.29 Mb. Скачать 3.29 Mb.
|

|
Тема: «ПНС» Вопрос 1. Строение периферического нервного ствола. Оболочки нерва: эндо-, пери- и эпиневрий. Периневральный барьер: структура, функции. Периферический нерв (нервный ствол) представляет собой пучок нервных волокон. В составе нерва имеются следующие оболочки: 1.Эндоневрий – представляет собой рыхлую соединительную ткань окружающую каждую нервное волокно по отдельности, и заполняющее промежутки между волокнами. 2.Периневрий состоит из наружной и внутренней частей. Наружная – плотная соединительная ткань окружающая каждый пучок нервных волокон. Внутренняя часть – несколько слоев периневральных клеток, образующих эпителиоподобный пласт, соединенных плотными контактами, покрытых толстой базальной мембраной снаружи и изнутри. Внутренняя часть периневрия образует периневральный барьер, контролирующий транспорт молекул к нервным волокнам и защищающая нервные волокна от повреждений, а также предотвращает проникновение в эндоневрий инфекционных агентов. 3.Эпиневрий – волокнистая соединительная ткань, объединяющая все пучки в составе нерва. В состав пери- и эпиневрия входят артериолы и венулы, а в состав эндоневрия – кровеносные капилляры. Во всех трех слоях присутствуют nervi nervorum, которые являются ветвями самого нерва. Вопрос 2. Локализация, строение и функции шванновских клеток. Миелинобразующие и миелиннеобразующие терминальные шванновские клетки. Роль шванновских клеток в регенерации периферических нервных проводников. Шванновские клетки входят в состав миелиновых и безмиелиновых периферических нервых волокон, синтезируют белки Р0, Р1, Р2, охватывают нервные волокна: в безмиелиновых они образуют мезаксон, а в миелиновых мембрана шванновской клетки концентрически наслаивается на нервное волокно, образуя миелин. Шванновские клетки между собой образуют щелевые контакты. При повреждении нервного волокна в составе периферического нерва, шванновские клетки принимают участие в его регенерации, образуя бюнгнеровские ленты – направляющие пути для регенерации аксонов. Шванновские клетки в составе бюнгнеровских лент синтезируют и секретируют нейротрофические факторы и рецепторы к ним. Те шванновские клетки до которых дорос регенерирующий аксон прекращают синтез нейротрофических факторов. Вопрос 3. Миелиновые и безмиелиновые волокна, их строение. Узловые перехваты. Миелиновые насечки. В зависимости от того, формируют ли шванновские клетки вокруг осевого цилиндра миелин, выделяют безмиелиновые и миелинизированные нервные волокна. В безмиелиновых нервных волокнах осевой цилиндр погружается в шванновскую клетку, мембрана которой смыкается, образуя мезаксон – сдвоенная мембрана шванновской клетки. В миелинизированном нервном волокне мембрана мезаксона удлиняется и концентрически наслаивается на волокно, мембранны в составе миелина расслаиваются, образуя насечки Шмидта-Лантермана, в которых присутствует цитоплазма шванновских клеток. Строение миелинового нервного волокна. Каждая шванновския клетка миелинизирует небольшой участок аксона, снаружи от миелина располагаются тонкий слой цитоплазмы и ядро шванновской клетки. Миелин прерывается через регулярнае промежутки – перехваты Ранвье. Мембрана аксонов в районе перехватов Ранвье содержит много натриевых каналов, необходимых для поддержания импульсной активности. Вопрос 4. Дегенерация и регенерация нервных волокон в периферическом нерве. После нарушения целостности нервного волокна происходит тигролиз (распыление вещества Ниссля), что означает прекращение синтеза белка в перикарионе и аксонного транспорта. При повреждении дегенереация нервного волокна происходит на небольшом протяжении центрального и на всем протяжении периферичексого отрезка – Уоллеровская дегенерация. Остатки дегенерировавших участков (остатки разрушенного осевого цилиндра и распавшегося миелина) аксона фагоцитируются макрофагами и частично шванновскими клетками. Шванновские клетки, утратившие связь с аксоном формируют Бюнгнеровские ленты и начинают продуцировать фактор роста нервов и рецепторы к нему же. Шванновской клеткой выделяются различные медиаторы, которые поглощаются аксоном, транспортируются в перикарион, где стимулируют синтез белка. Конус роста аксона перемещается по Бюнгнеровским лентам, отслаивая базальную мембрану, покрывающую шваннвскую клетку. Те шванновские клетки, до которых дорос регенерирующий аксон, прекращают синтез факторов роста нервов. Скорость роста составляет 0, 25 мм/сут, а после прохождения зоны травмы - 3-4 мм/сут. Вопрос 5. Строение спинномозгового узла. Морфофункциональная характеристика псевдоуниполярных нейронов, клеток-сателлитов. Спинномозговой узел имеет веретеновидную форму и покрыт капсулой из плотной волокнистой соединительной ткани. Внутри узла находятся группы нервных клеток, между которыми проходят пучки миелиновых нервных волокон. Чувствительные нервные клетки узла имеют вырост, который на расстоянии от округлого тела клетки Т-образно ветвится. В теле нейрона видны светлое крупное ядро с центрально расположенным ядрышком, в перикарионе — многочисленные глыбки хроматофильного вещества (Ниссля) (при окрашивании основным красителем). Каждый нейрон окружён небольшими клетками-сателлитами, которые имеют уплощённое тело с длинными иногда ветвящимися отростками. Они обладают, как и прочие клетки глии, способностью к фагоцитозу (они внедряются в тело нервной клетки и разрушают его, образуя предварительно ямки на его поверхности). Возможно также, что они служат для питания нервных элементов. Вопрос 6. Классификация двигательных и чувствительных нервных окончаний. Чувствительные окончания классифицируются на: свободные, несвободные, инкапсулированные. Свободные - терминальные ветвления периферического отростка чувствительного нейрона. К несвободным относится комплекс тельца Меркеля с нервной терминалью. К инкапсулированным: тельце Пачини, тельце Мейсснера, сухожильный орган Гольджи, тельце Руффини, мышечные веретена. К двигательным относятся: нервно-мышечный синапс и двигательные нервные окончания среди ГМК. Вопрос 7. Чувствительные нервные окончания: структурно-функциональная классификация, локализация, функции. Чувствительные нервные окончания делятся на: 1.Свободные нервные окончания – терминальные ветвления периферического отростка чувствительного нейрона. Они расположены в прослойках соединительной ткани внутренних органов, в сосочках кожи, в базальном и шиповатом слоях эпидермиса. Осевой цилиндр контактирует с вспомогательными клетками на значительном протяжении или окружен ими. В центре расположена эллиптической формы нервная терминаль, а снаружи вспомогательные клетки. Функции: регистрируют изменения температуры, рН, рО2, рСО2. 2.Несвободные нервные окончания – нервные окончания, содержащие в своём составе, кроме терминальных ветвлений периферического отростка, специальные клетки. К ним относятся Это преимущественно механорецепторы, которые локализованы в эпидермисе на базальной мембране, в соединительной ткани кожи и суставов, а также в мышцах. К ним относится комплекс клетки меркеля с нервной терминалью. 3.Инкапсулированные нервные окончания – несвободные окончания, имеющие оформленную соединительнотканную капсулу. К ним относятся: тельце Пачини, тельце Мейсснера, сухожильный орган Гольджи, тельце Руффини, мышечные веретена. Вопрос 8. Свободные нервные окончания: строение, функции. Они расположены в прослойках соединительной ткани внутренних органов, в сосочках кожи, в базальном и шиповатом слоях эпидермиса. На самом деле они не свободны, т. к. терминали как правило окружены вспомогательными клетками – аналогами Шванновских клеток. Осевой цилиндр контактирует с вспомогательными клетками на значительном протяжении или окружен ими. В центре расположена эллиптической формы нервная терминаль, а снаружи вспомогательные клетки. Терминаль может образовывать пальцевидные выросты между вспомогательными клетками. Функции: регистрируют изменения температуры, рН, рО2, рСО2. Вопрос 9. Комплекс осязательного эпителиоцита (клетки Меркеля) с нервной терминалью. Строение, локализация, функция. Комплекс клетки Меркеля с нервной терминалью образован несвободным нервным окончанием и клеткой Меркеля. Локализованы на базальной мембране в эпидермисе. Клетки Меркеля – округлые удлиненные клетки, формируют контакт с нервной терминалью чувствительного нервного волокна, они соединены с кератиноцитами при помощи десмосом, образуя пальцевидные выпячивания. В части клетки, обращенной к нервной терминали локализвано некоторое количество специфических осмиофильных гранул. Функции: являются осязательными механорецепторами, могут нести информацию об окончательной локализации для нервной терминали. Вопрос 10 Веретеновидные тельца. Строение, локализация, функции. Крупные рецепторы веретеновидной формы длиной до 2 мм и диаметром 150 мкм, расположены в соединительной ткани кожи суставов. Снаружи тельце покрыто соединительнотканной капсулой из нескольких слоев уплощенных фибробластов. Сердцевину рецептора образуют ветрвящиеся нервные терминали, покрытые базальной мембраной и окруженные (но не покрытые!) вспомогательными клетками, аналогами шванновских. Между соединительнотканной капсулой и нервной терминалью расположено заполненное жидкостью капсулярное пространство, которое содержит фибробласты, макрофаги и неорийнтированные коллагеновые волокна. Является механорецептором. Вопрос 11. Пластинчатые тельца (Пачини). Строение, локализация, функции. Расположены в соединительной ткани различных органов, в коже – в подкожной клетчатке. Образованы нервной терминалью, внутренней колбой и наружной капсулой. К наружной капсуле подходит миелинезированое нервное волокно, в наружной капсуле оно еще образует несколько перехватов Ранвье, после чего совсем теряет миелин и переходит во внутреннюю капсулу. Центральную часть внутренней колбы занимает одиночное безмиелиновое волокно. Внутренняя колба состоит из нескольких десятков отростчатых клеток, образующих 2 разделенные щелями полуокружности. По периферии внутренней колбы расположены тела ее клеток. Наружная капсула образована множеством уплощенных фибробластов. Функции: является механорецептором, наружная капсула рецептора является фильтром, пропускающим только динамическую составляющую механического воздействия. Вопрос 12. Тактильные тельца (Мейсснера). Строение, локализация, функция. Присутствуют в сосочковом слое кожи. Имеет удлиненную форму, снаружи покрыто соединительнотканной капсулой, в которую вплетаются коллагеновые волокна окружающей соединительной ткани. В формировании тельца участвуют несколько миелиновых волокон, а подходя к тельцу они теряют миелин. Сердцевина тельца образована пластинчатыми вспомогательными клетками, чередующимися с расширенными нервными терминалями. Являются механорецепторами. Вопрос 13. Мышечные веретена. Строение, локализация, функция. Расположены в скелетной мышце. Мышечное веретено содержит несколько коротких интрафузальных мышечных волокон, которые делятся на волокна с ядерной сумкой (ядра образуют компактные скопления в средней части интрафузального мышечного волокна) и на волокна с ядерной цепочкой (ядра распределены по всей длине). К мышечным волокнам подходят нервные волокна трех видов. Терминали Iα-волокон (чувствительных), спиралевидно окружают мышечные волокна обоих видов в середине. Терминали волокон II типа (чувствительных) разветвляются на мышечных волокнах в области рядом с серединой. Терминали Aγ-волокон (двигательных) образуют синапсы с мышечными волокнами в концевой их части. Само веретено окружено многослойной капсулой, наружный слой которой является производным периневрия, а внутренний эндоневрия нервного ствола, терминали волокон которого разветвляются на мышечных волокнах внутри тельца. Функции: являются рецепторами напряжения. Вопрос 14. Сухожильные органы (Гольджи). Строение, локализация, функция. Локализован в сухожилии на границе его с мышцей, а также в связках капсулы суставов. Имеет веретеновидную форму и окружен соединительнотканной капсулой – производной периневрия нервного пучка, волокна которого подходят к капсуле. Капсула содержит капилляры. Терминали афферентных миелиновых волокон ветвятся среди пучков коллагеновых волокон в сухожилии. Полость капсулы заполнена жидкостью. Сухожильный орган Гольджи является рецептором растяжения мышц. Вопрос 15. Двигательные нервные окончания. Строения, локализация, функции. Двигательные нервные окончания представлены нервно-мышечными синапсами и двигательными нервными окончаниями среди ГМК. Нервно-мышечный синапс состоит из пресинаптической мембраны (специализированная часть аксолеммы нервной терминали), в которой локализованы синаптические пузырьки, образующиеся в перикарионе, содержащие медиатор ацетилхолин, синаптической щели и постсинаптической мембраны, образующей многочисленные инвагинации и имеющей на своей поверхности особые белки – никотиновые холинорецепторы. Кроме того, имеется синаптическая базальная мембрана, проходящая через синаптическую щель. Она удерживает в области синапса терминаль аксона, контролирует расположение холинорецепторов в виде скоплений в постсинаптической мембране. Активация кальциевых каналов пресинаптической мембраны способствует высвобождению синаптических пузырьков, которые, сливаясь с пресинаптической мембраной, высвобождают в синаптическую щель ацетилхолин. Он связывается с холинорецепторами на постсинаптической мембране, активируя ионные каналы, через которые проходят потенциал-зависимые ионы (ионы натрия, хлора, калия и кальция), генерируя в постсинаптической мембране ПД, который распространяется по мышце. Двигательная иннервация ГМК отличается тем, что нервные волокна образуют на своём протяжении варикозные расширения, в которых локализованы синаптические пузырьки с нейромедиаторами (норадреналин и ацетилхолин), которые диффундируют из варикозных расширений в межклеточное пространство и взаимодействуют с их рецепторами в плазмолемме ГМК. В варикозных расширениях также локализованы нейротрансмиттеры, которые усиливают или угнетают действие основного медиатора. При секреции норадреналина симпатическими (адренергическими) волокнами вызывается сокращение ГМК, а при секреции ацетилхолина парасимпатическими (холинергическими) волокнами – их расслабление. Как правило, в составе гладких мышц, иннервированы не все ГМК; возбуждение неиннервированных ГМК происходит либо при медленной диффузии медиатора, либо посредством проведения потенциала действия через щелевые контакты между гладкомышечными клетками. Вопрос 16. Нервно-мышечный синапс. Строение, локализация, функции. Нервно-мышечный синапс состоит из пресинаптической мембраны (специализированная часть аксолеммы нервной терминали), в которой локализованы синаптические пузырьки, образующиеся в перикарионе, содержащие медиатор ацетилхолин, синаптической щели и постсинаптической мембраны, образующей многочисленные инвагинации и имеющей на своей поверхности особые белки – никотиновые холинорецепторы. Кроме того, имеется синаптическая базальная мембрана, проходящая через синаптическую щель. Она удерживает в области синапса терминаль аксона, контролирует расположение холинорецепторов в виде скоплений в постсинаптической мембране. Активация кальциевых каналов пресинаптической мембраны способствует высвобождению синаптических пузырьков, которые, сливаясь с пресинаптической мембраной, высвобождают в синаптическую щель ацетилхолин. Он связывается с холинорецепторами на постсинаптической мембране, активируя ионные каналы, через которые проходят потенциал-зависимые ионы (ионы натрия, хлора, калия и кальция), генерируя в постсинаптической мембране ПД, который распространяется по мышце. Вопрос 17. Вегетативные ганглии. Строение, локализация, функции. Синапсы вегетативных нейронов: варикозные расширения, нейромедиаторы. Капсула вегетативного ганглия покрывает его снаружи, образована рыхлой соединительной тканью, прослойки которой вдаются внутрь органа. Нервные клетки располагаются диффузно, все нейроны мультиполярные, двигательные. Нейромедиаторами в симпатической нервной системе являются норадриналин, в парасимпатической – ацетилхолин. От нервных клеток отходят отростки, дендриты и аксоны, которые вместе с оболочками образуют нервные волокна, которые естественно имеются в ганглиях. В симпатических ганглиях вегетативной нервной системы имеются МИФ-клетки (малые интенсивно-флуоресцирующие клетки) - небольших размеров нервные клетки, содержат серотонин, регулируют проведение импульсов с преганглионарных волокон на нейроны ганглия, от которых отходят постганглионарные волокна В парасимпатических ганглиях вегетативной нервной системы нервные клетки разделяются на 3 типа: - Клетки Догеля I типа - постганглионарные эфферентные нейроны; - Клетки Догеля II типа - чувствительные нейроны местных рефлекторных дуг, образуют синапсы с клетками I типа; - Клетки Догеля III типа - ассоциативные нейроны, связывающие соседние ганглии. Тема 11: «Центральная нервная система» Вопрос 1. Нервная трубка как источник развития головного и спинного мозга, слои нервной трубки. Радиальная глия, интекинетическая миграция ядер. Головной отдел образует 3 расширения (мозговых пузыря): 1) Передний мозг, на 5 неделе разделятся: - конечный мозг, образован первичными полусферами мозга, являются боковыми желудочками; - промежуточный мозг с глазными пузырями, полость является 3 желудочком. 2) Средний мозг, от ромбовидного мозга(3) отделен глубоким перешейком. 3) Ромбовидный мозг, разделяется: - задний мозг. Развивается мост и мозжечок, полость мозга является 4 желудочком; - миелэнцефалон. Просвет между 3 и 4 желудочком сужается и образуется сильвиев водопровод, боковые желудочки сообщаются через внутрижелудочковые отверстия Монро. Остальная часть нервной трубки формирует спинной мозг. Радиальная глия - глиальные клетки с длинными отростками, играющие важную роль в нейрональной миграции, построении слоёв коры мозга и мозжечка, а также являющиеся предшественниками в процессе нейрогенеза. Из этого запаса образуются нейроциты-основные клетки нервной ткани. Вопрос 2. Серое вещество спинного мозга (пластинки, ядра). Образовано из отростков и перикарионов, образующих скопления – ядра, объединённые в пластинки. Топография ядер, как правило, соответствует топографии пластинок. Всего существует 10 пластинок: Пластинка 1. Слой серого вещества у поверхности заднего столба, содержит различные по размеру нейроны. Перикарионы нейронов образуют контакты с аксонами из пластинки 2, а аксоны псевдоуниполярных нейронов оканчиваются на дендритах части нейронов пластинки 1. Нейроны данной пластинки реагируют на болевые и температурные стимулы и отдают волокна спиноталамическому пути противоположной стороны. Присутствуют нейроны содержащие энкефалин; волокна дающие положительную иммуноцитохимическую реакцию на вещество Р, энкефалин, соматостатин и серотонин. Пластинка 2. Располагается вентромедиально от пластинки 1, содержит скопление мелких нейронов – роландово студенистое вещество. Входят нейроны из краевого пояса Лиссауэра, и из частей бокового канатика. К нейронам внутренней зоны пластинки подходят волокна болевой и температурной чувствительности, к внешней зоне волокна тактильной чувствительности. Нейроны пластинки влияют на возбудимость крупных нейронов центральных отделов серого вещества. Так же содержит вещество Р, нейромедиатор передачи возбуждения от терминалей центральных отростков псевдоуниполярных нейронов спинномозговых узлов к вставочным нейронам спинного мозга, и энкефалин. Пластинка 3. Ветви дендритов оканчиваются в пластинках 1 и 2, аксоны делятся образуя сплетение в пластинках 3 и 4. Пластинка 4. Тактильная чувствительность, некоторые нейроны проходят через переднюю белую спайку на противоположную сторону и идут к таламусу. Пластинка 5. Расположена в основании заднего рога. Латеральная часть участвует в образовании ретикулярной формации. Некоторые дендриты направляются в пластинку 2. Пластинка 6. Присутствует в утолщениях спинного мозга. В медиальной части – окончание мышечных афферентов, в латеральной – нисходящие спинальные пути. Пластинка 7. Расположена между передними и задними рогами. Одни нейроны вместе с другими нейронами пластинок 5 и 6 образуют перекрещивающиеся волокна переднего спиномозжечкового пути, а другие образуют несколько ядер. Ядра: - Дорсальное ядро, Кларка (nucleus dorsalis). Образовано нейронами расположенными медиально, C8-L2. Аксоны образуют задний спиномозжечковый путь (Флексига). - Промежуточное боковое ядро (nucleus intermediolateralis). Расположено Th1 – L2. Нейроны дают начало преганглионарным симпатическим волокнам. - Промежуточное медиальное ядро (nucleus intermediomedialis). Расположено в медиальной части, латеральнее центрального канала. Получает висцеральные афферентны на всех уровнях спинного мозга. - Центральное шейное ядро. Скопление нейронов в верхних шейных сегментах, латеральнее промежуточного медиального. Аксоны, перекрещиваясь, проходят в мозжечок и нижнее вестибулярное ядро. Пластинка 8. В утолщениях пластинка в средней части передних рогов, в остальных частях в основании рогов, вентральнее пластинки 7. Пластинка 9. Содержит скопления двигательных нейронов. Различают медиальную и латеральную группу ядер. Медиальная по всей длине и в ней имеются переднемедиальные и заднемедиальные ядра. Иннервируют мускулатуру туловища. Остальные скелетные мышцы иннервируются латеральной группой. Пластинка 10. Область серого вещества, прилежащая к центральному каналу. Вопрос 3. Нейроны спинного мозга: двигательные, вегетативные, вставочные. Локализация, морфо-функциональная характеристика. Двигательные нейроны: - альфа-мотонейроны, входят в состав медиальных и латеральных ядер. Аксоны образуют нервно-мышечные синапсы и участвуют в формировании нейромоторных единиц. Так же посылают коллатерали к вставочным нейронам, образующим тормозные синапсы. - гамма-мотонейроны, иннервируют интрафузальные волокна мышечных веретен. Вставочные нейроны, передают информацию от одних нейронов к другим: -Клетки Реншоу регистрируют сигналы от альфа-мотонейронов, образуют тормозные синапсы. Вегетативные нейроны, расположены в висцеральных ядрах грудного и поясничного отделов (промежуточное боковое и промежуточное медиальное ядро), а так же в крестцовом отделе (ядро Онуфровича). Нейроны промежуточного медиального образуют синапсы с афферентными волокнами общей висцеральной чувствительности, а аксоны вступают в контакт с промежутным боковым ядром, а его клетки – преганглионарные симпатические нейроны. Ядро Онуфровича – преганглионарные парасимпатические нейроны. Вопрос 4. Белое вещество спинного мозга (проводящие пути). Морфологическая характеристика. Состоит из нерных волокон и клеток нейроглии. Разделено на три канатика: - Задние, между задней бороздой и задними корешками; - Боковые, между корешками; - Передние, между передней щелью и передними корешками. Кпереди от серой спайки – белая спайка, соединяет передние канатики. Проводящие пути – цепи нейронов, соединённые своими отростками, проводящие возбуждение. Вопрос 5. Восходящие чувствительные пути. Образованы центральными отростками чувствительных нейронов спинномозговых узлов и аксонами вставочных нейронов. Центральные остростки: - Тонкий пучок (Голля). Проходит в составе заднего канатика и заканчивается в тонком ядре продолговатого мозга. Проводящий путь проприоцептивной и тактильной чувствительности. - Клиновидный пучок (Бурдаха). Проходит в заднем канатике и заканчивается в клиновидном ядре продолговатого мозга. Проприоцептивной и тактильной чувствительности. Аксоны вставочных нейронов: - Вентральный спиноталамический путь. Проходит в переднем канатике отростки первых нейронов, расположенных в спинномозговых узлах, проводят тактильные и прессорные ощущения от механорецепторов кожи. Центральные отростки вступают в задние канатики, поднимаются и образуют синапсы с нейронами задних рогов. Далее аксоны переходят через переднюю спайку на противоположную сторону и восходят к заднелатеральному вентральному ядру таламуса. - Латеральный спиноталамический путь. Проходит в боковом канатике. Периферические окончания – свободные нервные окончания кожи. Центральные отростки спинномозговых узлов входят в спинной мозг через задние канатики и поднявшись на 1-2 сегмента, образуют синапсы с нейронами роландова студенистого вещества. Далее идут через переднюю спайку на противоположную сторону и поднимаются в боковых канатиках, заканчиваются в вентролатеральных ядрах таламуса. Главный путь болевой и температурной чувствительности. - Передний спинно-мозжечковый путь. Проприоцептивный путь мозжечка, проходящий в латеральном канатике, перекрещивается и вступает в верхнюю ножку мозжечка противоположной стороны и заканчивается в коре червя мозжечка. - Задний спинно-мозжечковый путь (Флексига). Проприоцептивный путь мозжечка, проходящий в боковом канатике и через нижнюю ножку мозжечка в кору червя мозжечка. По заднему и переднему мозжечковым путям поступает информация о всех афферентных сигналах глубокой чувствительности и о всех изменениях мышечного тонуса (координация произвольных движений). - Спинооливный путь, пучок переднего канатика, заканчиваясь в добавочных ядрах оливы. - Спинопокрышечный путь, путь общей чувствительности. Проходит в переднем канатикеи заканчивается в нижнем и верхнем холмиках крыши среднего мозга противоположной стороны. - Спиноретикулярный путь – волокна бокового канатика, конец в ретикулярной формации продолговатого мозга, моста и среднего мозга. Получение информации соматических и висцеральных рефлексов. Вопрос 6. Нисходящие двигательные пути. Заканчиваются на мотонейронах. Супраспинальные: - Передний корково-спинномозговой путь (пирамидный путь передний). Образован аксонами нейронов в двигательной зоне коры предцентральной извилины. Волокна проходят через внутреннюю капсулу и в переднем канатике, заканчивается в передних рогах, перекрещиваясь. - Латеральный корково-спинномозговой путь (пирамидный латеральный). Начинается в коре предцентральной извилины, проходит через внутреннюю капсулу и проходит в боковой канатик, заканчиваясь в передних рогах. - Тектоспинальный путь. Начинается в верхних холмиках, проходит через передний канатик и заканчивается в передних рогах. - Красноядерно-спинномозговой путь (фон Монакова), путь экстрапирамидной системы. Начинается от красного ядра, проходит в боковом канатике и заканчивается в передних рогах. - Ретикулоспинальный путь. Путь экстрапирамидной системы. Начинается в ретикулярной формации, заканчивается в передних рогах. Контролирует тонус скелетной мускулатуры и висцеральные двигательные функции. - Преддверно-спинномозговой путь (Гельда). Латеральный начинается от латерального вестибулярного ядра (Дейтерса), проходит в переднем канатике и достигает в передних рогах альфа- и гамма-мотонейронов. Аксоны медиального ядра (Швальбе) присоединяются к медиальному продольному пучку и спускаются вниз в виде медиального преддверно-спиномозгового пути. - Оливоспинномозговой путь (Гельвега). От оливного ядра, проходят в переднем канатике и заканчивается в передних рогах. Спинальные: - В задних канатиках. Клетки ядер в задних столбах дают начало нисходящим путям в ипсилатеральных задних канатиках и заканчиваются в пластинках 4, 5, и возможно 1. Пучок в форме запятой (Шультце). Нисходящие ветви центральных отростков достигают дорсального ядра, а частично нейронов пластинки 6. В шейных и верхних грудных сегментах образуют пучок в форме запятой. Септомаргинальный пучок. Образован так же нисходящими волокнами. - В передних канатиках. Бороздчато-краевой пучок. Волокна медиального преддверно-спинномозгового пути, спускающийся вниз и оканчивающийся в передней части грудного отдела спинного мозга. Влияют на тонус мышц шеи. Вопрос 7. Кора мозжечка. Слои, нейроны, взаимосвязи между нейронами. Кора мозжечка (серое вещество) имеет три слоя: 1) Молекулярный слой (наружный). Содержит: - Корзинчатые клетки – мультиполярные нейроны. Образуют многочисленные длинные и сравнительно мало разветвленные дендриты. Аксон образует ветви, заканчивающиеся в виде корзинок на телах клеток Пуркинье. - Звёздчатые клетки расположены ближе к поверхности коры. Синапсы с дендритами клеток Пуркинье. 2) Ганглионарный слой (средний). Образуют тела клеток Пуркинье. Перикарионы имеют грушевидную форму и образуют пласт ровным слоем от поверхности коры. В молекулярный слой отходят 2-3 ветвящихся дендрита. Через зернистый слой в белое вещество от тела клетки Пуркинье отходит аксон. Вблизи тела клетки от аксона отходят коллатерали, направляющиеся обратно в ганглионарный слой и глубокие части молекулярного слоя, здесь они контактируют с другими клетками Пуркинье. На клетках Пуркинье заканчиваются все афферентные пути мозжечка. 3) Зернистый слой (внутренний). Содержит: - Клетки-зёрна. Невелики. 3-4 коротких дендриты образуют концевые разветвления (птичьи лапки). Аксоны поднимаются в молекулярный слой, где образуют разветвления, которые образуют синапсы. - Клетки Гольджи типа 2. Аксоны клеток, входя в состав клубочков мозжечка, заканчиваются синаптическими контактами на розетках моховидных волокон. На них заканчивается часть коллатералей аксонов грушевидных клеток Пуркинье. Вопрос 8. Клубочки мозжечка. Клубочки мозжечка – скопление терминальных ветвлений отростков различных нейронов мозжечка и моховидных волокон. Клубочек окружен капсулой из глиальных клеток. Вокруг расположены скопления клеток-зёрен. Содержит: розетки – окончания моховидных волокон; птичьи лапки; аксоны и часть разветвления дендритов клеток Гольджи типа 2. Вопрос 9. Слои и нейроны коры головного мозга. Слои коры (снаружи внутрь): Молекулярный. Содержит редкие перикарионы, проходят аксоны и дендриты. Наружный зернистый. Присутствуют небольшие пирамидные и звёдчатые нейроны. Наружный пирамидный. Многочисленные пирамидные нейроны средней величины. Внутренний зернистый. Мелкие звёздчатые клетки. Внутренний пирамидный (гнаглионарный). Крупные пирамидные и небольшое количество звёздчатых нейронов. Полиморфный. Множество нейронов различной величины, а также пирамидных и зернистых нейронов. Нейроны: - Пирамидные нейроны. От основания пирамиды отходит аксон, уходящий в белое вещество. Возвратные коллатеральные ветви аксона заканчиваются на других пирамидных нейронах или вставочных корковых нейронах. Разновидности: А. Клетки Беца. Расположены в слое 5 двигательной коры, самые крупные. Б. Клетки Мейнерта. Крупные, расположены в слое 5 зрительной коры затылочной доли, они посылают аксоны в ствол мозга и участвуют в рефлексе движения глаза. - Звёздчатые нейроны. Аксон и дендриты отходят и участвуют в образовании внутрикорковых связей. - Веретеновидные нейроны. Чаще в слое 6. Аксон уходит глубоко в белое вещество. - Клетки Мартинотти. Во всех слоях, кроме первого. Аксон отдает коллатерали во всех слоях. - Горизонтальные нейроны Рамон-и-Кахала. В первом слое. Отходит длинный аксон, который вместе с дендритами образует горизонтальные связи в пределах слоя 1. Вопрос 10. Желудочки мозга: хориоэпителиальная пластинка, строение и функции. Желудочки головного мозга - система анастомозирующих полостей, сообщающихся с центральным каналом спинного мозга и субарахноидальным пространством, содержащих спинномозговую жидкость и выстланных однослойным пластом клеток эпендимой глии низкопризматической или кубической формы с микроворсинками и ресничками на апикальной поверхности. В отдельных участках эпендимоциты обладают специфическими структурно-функциональными особенностями и принимают участие в выработке спинномозговой жидкости и химической сигнализации. Сосудистые сплетения желудочков головного мозга - структуры в области крыши III и IV желудочков, а также части стенок боковых желудочков, которые обеспечивают выработку 70-90 % спинномозговой жидкости (10-30 % вырабатываются тканями центральной нервной системы и выделяются эпендимой вне области сосудистых сплетений). Они образованы ветвящимися выпячиваниями мягкой мозговой оболочки, которые вдаются в просвет желудочков и покрыты особыми кубическими хороидными эпендимоцитами. Хороидные эпендимоциты содержат большое количество митохондрий, умеренно развитый синтетический аппарат, многочисленные пузырьки и лизосомы. Их выпуклая апикальная поверхность покрыта многочисленными микроворсинками. Латеральные поверхности формируют интердигитации и связаны комплексами соединений. Базальная поверхность образует переплетающиеся выросты (базальный лабиринт). По поверхности эпендимы сосудистых сплетений перемещаются уплощенные отростчатые клетки Кольмера с хорошо развитым лизосомальным аппаратом, которые, очевидно, являются макрофагами. Слой эпендимоцитов располагается на базальной мембране, отделяющей его от подлежащей рыхлой волокнистой соединительной ткани мягкой мозговой оболочки, в которой находятся многочисленные фенестрированные капилляры и встречаются слоистые обызвествленные тельца. Избирательная ультрафильтрация компонентов плазмы крови с образованием спинномозговой жидкости происходит из капилляров в просвет желудочков через гемато-ликворный барьер. Установлено, что клетки эпендимы способны также секретировать некоторые белки в спинномозговой жидкости и частично поглощать вещества из спинномозговой жидкости (очищая ее от продуктов метаболизма мозга, лекарств, в частности, антибиотиков). Гемато-ликворный барьер включает в себя: цитоплазму фенестрированных эндотелиальных клеток капилляров; базальную мембрану эндотелия капилляров; перикапиллярное пространство - широкое, содержащее рыхлую волокнистую соединительную ткань мягкой мозговой оболочки с большим количеством макрофагов; базальную мембрану эпендимы; слой хороидных эпендимных клеток. Спинномозговая жидкость циркулирует в субарахноидальном пространстве желудочках головного мозга и центральном канале спинного мозга. Ее общий объем у взрослого составляет 140-150 мл. Она вырабатывается в количестве 500 мл в сутки, полностью обновляясь каждые 4-7 ч и по составу отличается от сыворотки крови - в ней резко снижено содержание белка и повышены концентрации натрия, калия и хлора. Спинномозговая жидкость содержит отдельные лимфоциты (не более 5 клеток на 1 мл). Всасывание компонентов спинномозговой жидкости в кровь происходит в области ворсинок паутинного сплетения, вдающихся в расширенные субдуральные пространства по средней линии головного мозга; в незначительной части оно осуществляется эпендимой сосудистых сплетений. Танициты - специализированные клетки эпендимы в латеральных участках стенки III желудочка, инфундибулярного кармана и срединного возвышения, которые обеспечивают связь между спинномозговой жидкостью в просвете желудочков мозга и кровью. Они имеют кубическую или призматическую форму, их апикальная поверхность покрыта микроворсинками и отдельными ресничками, а от базальной отходит длинный отросток, оканчивающийся пластинчатым расширением на кровеносном капилляре. Танициты поглощают из спинномозговой жидкости и транспортируют их по своему отростку в просвет сосудов. Вопрос 11. Оболочки мозга, их гистологическое строение. Все оболочки образованы волокнистой соединительной тканью. Мягкая мозговая оболочка, содержит переплетающиеся пучки коллагеновых волокон и сеть эластических волокон, множество равномерно распределённых кровеносных сосудов. Снаружи покрыта слоем плоских клеток. Паутинная оболочка. Сеть тонких соединительнотканных перегородок (трабекул), состоящих преимущественно из коллагеновых и эластических волокон. Оболочка изнутри и снаружи выстлана непрерывным слоем тонких уплощенных клеток. Пространство между трабекулами – субарахноидальное, заполненно цереброспинальной жидкостью. В местах где оболочка не прилегает вплотную к мозговой – цистерны, содержащие большое количество цереброспинальной жидкости. Твёрдая оболочка. Состоит из плотной волокнистой соединительной ткани, преобладают коллагеновые волокна. Пространство между твёрдой и паутинной – субдуральное. Содержит жидкость ( не цереброспинальную). Снаружи покрыта рыхлой соединительной тканью. Образует складки, формируя с надкостницей синусы. Вопрос 12. Вегетативная нервная система. Симпатический отдел. Парасимпатический отдел. Вегетативная нервная система иннервирует все внутренние органы, сердце и сосуды, экзокринные и эндокринные железы, органы чувств и подразделяется на 2 отдела - симпатический и парасимпатический. Каждый орган, как правило, получает и симпатическую, и парасимпатичеcкую иннервацию. 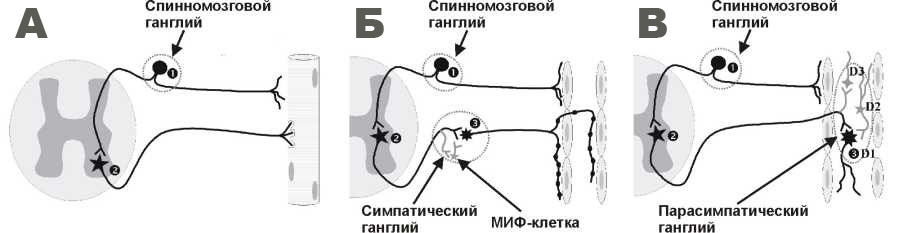 СИМПАТИЧЕСКАЯ НЕРВНАЯ СИСТЕМА Центры симпатической нервной системы находятся в боковых рогах грудного и поясничного отделов (CIII-LII или ThI-LIII) спинного мозга. Симпатическая рефлекторная дуга состоит как минимум из 3 нейронов: I нейрон - чувствительный, его перикарион лежит в спинномозговом ганглии, длинный дендрит отходит на периферию, где заканчивается рецептором, аксон входит в задние рога спинного мозга, проходит в боковой рог (или переключается на ассоциативный нейрон) и образует синапс со II нейроном; I нейрон - пурин-пептидергический, нейромедиаторы - АТФ, субстанция Р; II нейрон - называется преганглионарным; эфферентный, его перикарион и дендриты лежат в боковых рогах спинного мозга, аксон через передние рога выходит из спинного мозга и идет к симпатическому ганглию (нейроны узлов симпатического ствола или узлов периартериальных симпатических сплетений), где образует синапсы с III нейроном; II нейрон – холинергический, нейромедиатор – ацетилхолин; III нейрон - называется постганглионарным; эфферентный, его перикарион и дендриты лежат в симпатических ганглиях; на перикарионе и дендритах III нейрона имеются Н-холинорецепторы, через которые происходит синаптическая передача между II (преганглионарным) и III(постганглионарным) нейронами (ацетилхолин высвобождается из пресинаптической части, принадлежащей I нейрону и взаимодействует с Н-холинорецепторами, находящимися на постсинаптической мембране, то есть на мембране III нейрона). Аксон III нейрона выходит из ганглия и идет к иннервируемому органу, где образуются синаптические соединения. III нейрон - адренергический, нейромедиатор – норадреналин. Синаптическая передача между постганглионарным нейроном и рабочим органом осуществляется с помощью норадреналина, который высвобождается из пресинаптической части (принадлежащей III-нейрону) и взаимодействует с адренорецепторами, находящимися на постсинаптической мембране, которая принадлежит не нейрону, а органу. В различных органах имеются разные комбинации разных подтипов a-и b-адренорецепторов ПАРАСИМПАТИЧЕСКАЯ НЕРВНАЯ СИСТЕМА Центры парасимпатической нервной системы находятся в боковых рогах крестцового отдела спинного мозга (ScII-IV), продолговатом мозге и мосте (ядра III, VII, IX, X, черепномозговых нервов). Парасимпатическая рефлекторная дуга состоит как минимум из 3 нейронов: I нейрон - чувствительный, его перикарион лежит в спинномозговом ганглии или в толще нервного ствола или в специальных чувствительных ганглиях парасимпатичесокй нервной системы (ganglion geniculi, ganglia superius et inferius, g.nodosum), или непосредственно в стволе мозга (nucleus tractus solitarii - n. vagus; nucleus sensorius principallis nervi trigemini), длинный дендрит отходит на периферию, где заканчивается рецептором. Аксон входит в мозг (мост, продолговатый мозг), или в боковые рога спинного мозга (или переключается на ассоциативный нейрон) и образует синапс со II нейроном; I нейрон - пyрино-пептидергический, нейромедиаторы - АТФ, субстанция Р, кальцитонин ген-родственный пептид; II нейрон - называется преганглионарным; эфферентный, его перикарион и дендриты лежат в боковых рогах крестцового отдела спинного мозга или продолговатом мозге, мосте, аксон выходит из спинного мозга или в составе черепно-мозговых нервов идет к парасимпатическому ганглию, где образует синапсы с III нейроном; II нейрон - холинергический, нейромедиатор – ацетилхолин; III нейрон - называется постганглионарным; эфферентный, его перикарион и дендриты лежат в парасимпатических ганглиях (вегетативные ганглии черепно-мозговых нервов /g. ciliary, g.oticum, g pterygopalatinum, g. submandibulare/ или внутриорганные ганглии (интрамуральные) ганглии). На перикарионе и дендритах III нейрона имеются Н-холинорецепторы, через которые происходит синаптическая передача между II (преганглионарным) и III (постганглионарным) нейронами (ацетилхолин высвобождается из пресинаптической части, принадлежащей II нейрону, и взаимодействует с Н-холинорецепторами, находящимися на постсинаптической мембране, то есть на мембране III нейрона); аксон выходит из ганглия и идет к иннервируемому органу или уже находится в органе, где образуются синаптические соединения. III нейрон - холинергический, нейромедиатор – ацетилхолин. Синаптическая передача между постганглионарным нейроном и рабочим органом осуществляется с помощью ацетилхолина, который высвобождается из пресинаптической части и взаимодействует с М-холинорецепторами, находящимися на постсинаптической мембране синапса, а постсинаптическая мембрана - это уже мембрана не нейрона, а органа. |