Гистология реферат. Тема 1 клетка Вопрос Химический состав, организация плазмолеммы. Функции плазмолеммы
 Скачать 3.29 Mb. Скачать 3.29 Mb.
|

|
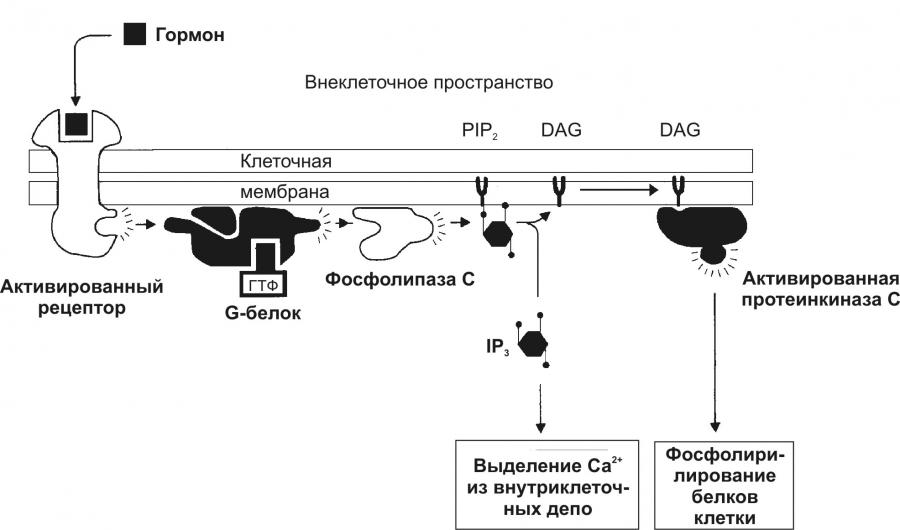
Тема 14: «Эндокринная система» Вопрос 1. Общая характеристика и классификация гормонов. Виды рецепторов, связывающих гормоны. Гормон – любое биологически активное вещество, циркулирующее во внутренней среде организма и оказывающее регуляторный эффект на свои клетки-мишени. Для гормонов характерна специфичность действия на конкретные клетки или органы, называемые мишенями, что обусловлено наличием на клетках-мишенях специфических рецепторов, распознающих и связывающих данный гормон. Будучи связанным с рецептором, гормон может воздействовать на плазматическую мембрану, на фермент, находящийся в этой мембране, на клеточные органеллы или на ядерный генетический материал. Рецептор – высокомолекулярное вещество, специфически связывающее конкретный лиганд (гормон). Выделяют 2 класса рецепторов, связывающих гормоны: Мембранные рецепторы (для пептидных гормонов), расположенные в плазматической мембране клетки. Реализация эффекта гормона, имеющего мембранные рецепторы, осуществляется с помощью второго посредника (цАМФ, цГМФ, ДАГ, ИТФ и др.). Общий механизм взаимодействия с клеткой пептидных гормонов:
Ядерные рецепторы (производные тирозина и ретиноевой кислоты, рецепторы стероидных гормонов), имеющие внутриклеточную локализацию. Общий механизм взаимодействия с клеткой стероидных гормонов:
Классификация гормонов по их мишеням: Рилизинг-гормоны – группа гормонов, синтезируемых в нейронах гипоталамуса, мишенями которых являются эндокринные клетки передней доли гипофиза. Рилизинг-гормоны подразделяются на: Либерины – рилизинг-гормоны, способствующие усилению синтеза и секреции соответствующих гормонов в эндокринных клетках передней доли гипофиза; Статины – рилизинг-гормоны, ингибирующие синтез и секрецию гормонов в клетках-мишенях гипофиза. Тропные гормоны – гормоны, клетками-мишенями которых являются другие эндокринные клетки. Тропные гормоны вырабатываются в передней доле гипофиза под влиянием рилизинг-гормонов гипоталамуса; Гормоны периферических эндокринных желёз, вырабатывающиеся под влиянием тропных гормонов передней доли гипофиза, непосредственно участвуют в регуляции функций организма. Классификация гормонов по химической природе: Гормоны-белки и гликопротеиды, рецепторы к которым располагаются на мембранах клеток-мишеней и, как правило, запускают систему вторичных посредников; Гормоны-производные аминокислот: йодсодержащие гормоны щитовидной железы, гормоны мозгового вещества надпочечников, гормоны эпифиза. Рецепторы к таким гормонам находятся в ядре, следовательно, данные гормоны способны непосредственно влиять на экспрессию гена; Гормоны-стероиды: гормоны коры надпочечников, половых желёз и гормон-витамин D3. Рецепторы к таким гормонам находятся в ядре, следовательно, данные гормоны способны непосредственно влиять на экспрессию гена. Вопрос 2. Особенности строения гормонпродуцирующих клеток. Эндокринная клетка – это клетка, синтезирующая и секретирующая во внутреннюю среду организма тот или иной гормон. Этот термин применим к клеткам желёз внутренней секреции, одиночным эндокринным клеткам и их небольшим скоплениям, часто объединяемым в диффузные эндокринные системы (например, энтеральная эндокринная система). Для эндокринных клеток, синтезирующих пептиды и белки, характерно наличие гранулярной ЭПС, где происходит сборка пептидной цепи, комплекса Гольджи, где происходит присоединение углеводных остатков, а также формирование секреторных гранул. Для эндокринных клеток, синтезирующих стероидные гормоны, характерно наличие развитой гладкой ЭПС, одной из функций которой является как раз синтез липидов и стероидных гормонов, и многочисленных митохондрий, вырабатывающих для этого необходимое количество энергии. Важным фактом является то, что эндокринные клетки находятся в тесном контакте с кровеносными капиллярами фенестрированного типа, имеющими широкий просвет. Вопрос 3. Общая характеристика эндокринных желёз, их отличия от экзокринных. К органам эндокринной системы относят: Центральные эндокринные органы: гипоталамус, гипофиз, эпифиз; Периферические эндокринные железы: щитовидная, паращитовидная железы и надпочечники (корковое и мозговое вещество); Органы, объединяющие эндокринные и неэндокринные функции: поджелудочная железа, гонады (семенники, яичники), плацента; а также вырабатывающие ряд гормоноподобных веществ почки и тимус. Одиночные гормонпродуцирующие клетки нервной, пищеварительной и дыхательной систем. Общая характеристика. Зрелые эндокринные железы не имеют выводных протоков и выделяют вырабатываемые ими гормоны во внутреннюю среду организма. Некоторые гормоны вырабатываются не отдельными железами, а группами компактно расположенных клеток (панкреатические островки, интерстициальные эндокриноциты яичка) или клеток, расположенных в различных органах и тканях (клетки APUD-системы). Эндокринные железы отличаюся от экзокринных: Эндокринные железы действуют дистантно. Секрет (гормон) эндокринных желез специфичен. Секрет (гормон) эндокринных желез (гормон) имеет высокую биологическую активность. Секрет (гормон) выделяется в ничтожно малых количествах. Развитые (зрелые, сформированные) эндокринные железы не имеют выводных протоков. Гормоны контролируют жизнедеятельность организма, все функции клеток, активность генов, формирование клеточного фенотипа. Регулируя активность ферментов, гормоны влияют на метаболизм. Все многообразие действия гормонов можно свести к трем важнейшим функциям: обеспечение роста и развития организма, обеспечение адаптации организма к постоянно меняющимся условиям среды и обеспечение гомеостаза. Вопрос 4. Эндокринная, паракринная и аутокринная типы регуляции. Различают три типа действия биоактивных веществ: Эндокринная (дистантная) регуляция – секреция гормона происходит во внутреннюю среду организма, клетки-мишени находятся далеко от эндокринной клетки. Пример: действие тропных гормонов на клетки-мишени. Паракринная регуляция – продуцент биоактивного вещества и клетка-мишень расположены рядом, молекулы гормона достигают мишени путём диффузии в межклеточном веществе. Пример: секрецию Н+ в париетальных клетках желёз желудка стимулируют гастрин и гистамин, а подавляют соматостатин и простагландины, секретируемые рядом расположенными клетками. Аутокринная регуляция – сама клетка-продуцент гормона имеет рецепторы к этому же гормону. Пример: В-лимфоциты секретируют фактор роста нервов и имеют к нему рецепторы, что является существенным для поддержания популяции В-клеток. Вопрос 5. Ядра и нейросекреторные клетки гипоталамуса. Гормоны гипоталамуса. Гипоталамус – высший нервный центр регуляции эндокринных функций, объединяющий эндокринные механизмы регуляции с нервными. Нейросекреторные клетки гипоталамуса – это нервные клетки гипоталамуса, в перикарионах которых синтезируется гормон и секретирующие этот гормон в кровь. Эти клетки получают афферентные нервные импульсы из других частей нервной системы. Аксоны же этих клеток оканчиваются на кровеносных сосудах, образуя аксо-вазальные синапсы, через которые в кровь выделяются гормоны. Для нейросекреторных клеток характерно наличие гранул нейросекрета. По ходу аксонов нейросекретоных нейронов гипоталамуса образуются утолщения этих аксонов – тельца Херинга, в которых накапливаются гранулы, содержащие гормоны. Непосредственно в терминалях аксонов сконцентрировано всего 30 % от общего количества гормонов, которое синтезируется в перикарионе данного аксона. В гипоталамусе условно выделяют передний, средний и задний отделы. В переднем отделе гипоталамуса располагаются парные супраоптические и паравентрикулярные ядра, в нейронах которых продуцируются вазопрессин (АДГ) и окситоцин соответственно. Вазопрессин (в сосудах), или антидиуретический гормон (в канальцах почек), вызывает усиление тонуса ГМК артериол, приводящее к повышению артериального давления, а также обеспечивает обратное всасывание жидкости, отфильтрованной в первичную мочу из крови. Окситоцин вызывает сокращение миометрия во время родов и сокращение миоэпителиальных клеток молочной железы. В среднем отделе гипоталамуса располагаются нейросекреторные ядра (Аркуатовентеромедиальная группа ядер), содержащие мелкие нейроны, вырабатывающие рилизинг-гормоны (либерины и статины), с помощью которых гипоталамус контролирует гормонобразовательную деятельность аденогипофиза. Либерины: соматолиберин, кортиколиберин, тиролиберин, гонадолиберин, меланолиберин, пролактолиберин. Статины: соматостатин, пролактоститин, меланостатин. Вопрос 6. Развитие и строение гипофиза. Анатомически гипофиз имеет ножку и тело, а гистологически подразделяется на аденогипофиз (передняя, туберальная и промежуторчная доли) и нейрогипофиз (задняя доля, нейрогипофизарная часть ножки и срединное возвышение). Гипофиз развивается из двух зачатков – эктодермального и нейрогенного. На 4-5 неделе эктодермальный эпителий образует карман Ратке – вырост крыши ротовой бухты, направляющийся к мозгу. Это образование даёт начало аденогипофизу. Навстречу карману Ратке растёт выпячивание промежуточного мозга, дающее начало нейрогипофизу (нейроэктодермальное происхождение). Гипофиз покрыт капсулой из плотной волокнистой соединительной ткани, его строма представлена тонкими прослойками соединительной ткани, связанными с сетью ретикулярных волокон, которые в аденогипофизе окружают тяжи эпителиальных клеток и мелкие сосуды. Передняя доля гипофиза образована разветвленными эпителиальными тяжами – трабекулами, промежутки между которыми заполнены рыхлой волокнистой соединительной тканью и капиллярами синусоидного типа, оплетающими трабекулы. Эндокриноциты, располагающиеся по периферии трабекул содержат в своей цитоплазме гранулы, интенсивно воспринимающие красители – хромофильные эндокриноциты (ацидофильные (соматотропы и лактотропы) и базофильные (гонадотропы, тиротропы и кортикотропы)). Клетки, занимающие середину трабекулы, имеют нечёткие границы, и их цитоплазма окрашивается слабо – хромофобные эндокриноциты (хромофильные клетки после выведения гранул секрета и малодифференцированные камбиальные элементы и фолликулярно-звёздчатые клетки). Промежуточная доля гипофиза представлена узкой полоской эпителия (у человека развита слабо), а ее эндокриноциты способны вырабатывать меланоцитстимулирующий и липотропный гормоны. Нейрогипофиз содержит отростки и терминали нейросекреторных клеток супраоптического и паравентрикулярного ядер гипоталамуса, по которым транспортируются и выделяются в кровь вазопрессин и окситоцин; многочисленные капилляры фенестрированного типа; клетки нейрогипофиза – питуициты – отростчатые глиальные клетки, выполняющие опорную и трофическую функцию. Вопрос 7. Развитие и строение аденогипофиза, клеточный состав. Аденогипофиз имеет эктодермальное происхождение. На 4-5 неделе эктодермальный эпителий образует карман Ратке – вырост крыши ротовой бухты, направляющийся к мозгу. Это образование даёт начало аденогипофизу. Аденогипофиз состоит из передней и промежуточной долей, а также туберальной части ножки гипофиза. Передняя доля гипофиза – эпителиальная эндокринная железа, клетки которой синтезируют и секретируют в кровь тропные гормоны. Она образована клетками, расположенными анастомозирующими тяжами и островками между прослойками рыхлой волокнистой соединительной ткани и кровеносными капиллярами фенестрированного типа (в которые выводятся тропные гормоны и из которых к эндокриноцитам поступают либерины и статины). Эндокринные клетки передней доли, синтезирующие пептидные гормоны, содержат элементы гранулярной ЭПС, комплекс Гольджи, много митохондрий и секреторных гранул. Эндокриноциты, располагающиеся по периферии эпителиальных тяжей и островков содержат в своей цитоплазме гранулы, интенсивно воспринимающие красители – хромофильные эндокриноциты, которые по окрашиванию секреторных гранул подразделяют на ацидофильные (соматотропы и лактотропы) и базофильные (гонадотропы, тиротропы и кортикотропы). Клетки, занимающие середину трабекулы, имеют нечёткие границы, и их цитоплазма окрашивается слабо – хромофобные эндокриноциты: дегранулировавшие хромофильные клетки, малодифференцированные камбиальные элементы и фолликулярно-звёздчатые клетки. Средняя доля гипофиза представлена узкой полоской эпителия (у человека выражена слабо), а ее эндокриноциты способны вырабатывать меланоцитстимулирующий и липотропный гормоны. Туберальная доля гипофиза состоит из тяжей эпителиальных клеток, между которыми расположены гипофизарные воротные вены, соединяющие первичную капиллярную сеть, находящуюся в районе срединного возвышения и вторичную капиллярную сеть, находящуюся в передней доле гипофиза. Эндокринная функция эпителиальных клеток туберальной доли практически отсутствует, здесь редко встречаются базофильные аденоциты. Вопрос 8. Развитие, строение, функции нейрогипофиза. Нейрогипофиз имеет нейроэктодермальное происхождение и развивается как выпячивание промежуточного мозга, растущее по направлению к карману Ратке – выросту крыши ротовой бухты, из которого развивается аденогипофиз. Нейрогипофиз состоит из задней доли гипофиза (здесь в кровь секретируются вазопрессин и окситоцин), нейрогипофизарной части ножки гипофиза (здесь в заднюю долю проходят аксоны гипоталамо-гипофизарного тракта) и срединного возвышения (здесь располагаются аксо-вазальные синапсы, в кровь поступают рилизинг-гомоны – первичная капиллярная сеть). Нейрогипофиз содержит: отростки и терминали нейросекреторных клеток супраоптического и паравентрикулярного ядер гипоталамуса, по которым транспортируются и выделяются в капилляры задней доли гипофиза вазопрессин и окситоцин; многочисленные капилляры фенестрированного типа; клетки нейрогипофиза – питуициты – отростчатые глиальные клетки, выполняющие опорную и трофическую функцию (эндокринной функции они не выполняют). Они содержат многочисленные промежуточные филаменты, пигментные гранулы и липидные включения. Вопрос 9. Гормоны аденогипофиза, их мишени и эффекты.
Вопрос 10. Гормоны нейрогипофиза, их мишени и эффекты. NB! В задней доле гипофиза гормоны не синтезируются – здесь лишь происходит поступление в кровь нейрогормонов (АДГ и окситоцина), образованных в гипоталамусе.
Вопрос 11. Система кровоснабжения гипофиза.
Вопрос 12. Рилизинг-гормоны гипоталамуса: их мишени и эффекты.
Вопрос 13. Гипоталамо-гипофизарный тракт: его организация, доставка гормонов, эффекты гормонов. Гипоталамо-гипофизарная система включает в себя: Переднюю долю гипофиза, где синтезируются тропные гормоны, мишенями которых являются другие эндокринные клетки; Перикарионы нейросекреторных нейронов гипоталамуса, в которых происходит синтез рилизинг-гормонов, регулирующих синтез и секрецию тропных гормонов (либеринов, увеличивающих синтез и секрецию, статинов – угнетающих их), а также вазопрессина (или АДГ), окситоцина и нейрофизинов. Необходимо отметить, что синтез рилизинг-гормонов происходит в диффузно расположенных нейронах гипоталамуса – в разных его областях, а синтез вазопрессина и окситоцина происходит в супраоптическом и паравентрикулярном ядрах, соответственно; Гипоталамо-гипофизарный тракт представлен аксонами нейросекреторных нейронов, по которым происходит транспорт гормонов из гипоталамуса в аденогипофиз (рилизинг-гормоны) и нейрогипофиз (АДГ, окситоцин). 70% таких гормонов содержится не непосредственно у нервной терминали, а по ходу аксона, образуя локальные расширения – так называемые тельца Херринга; Аксо-вазальные синапсы, образованные терминальными расширениями аксонов нейросекреторных нейронов гипоталамуса, контактирующими со стенкой кровеносных капилляров срединного возвышения (первичная капиллярная сеть) и задней доли гипофиза. Через капилляры срединного возвышения в кровь поступают рилизинг-гормоны, а в капилляры задней доли – вазопрессин и окситоцин; Портальную систему кровотока между срединным возвышением и передней долей гипофиза. Она состоит из: первичной капиллярной сети срединного возвышения, где заканчиваются терминали аксонов нейросекреторных клеток гипоталамуса; воротных вен туберальной части аденогипофиза, которые идут по гипофизарной ножке туберальной части в переднюю долю гипофиза; вторичной капиллярной сети передней доли, откуда обогащённая гормонами кровь поступает в общую циркуляцию через выносящие вены. Вопрос 14. Аксо-вазальные синапсы: локализация, структура, функции. Аксо-вазальные синапсы образованы терминальными расширениями аксонов нейросекреторных нейронов гипоталамуса, контактирующими со стенкой кровеносных капилляров срединного возвышения (здесь в кровь поступают рилизинг-гормоны) и задней доли гипофиза (здесь в кровь поступают АДГ и окситоцин), т.е.:
Вопрос 15. Эпифиз: строение, клеточный состав. Эпифиз, или шишковидная железа, - вырост промежуточного мозга, соединённый ножкой со стенкой III желудочка. Капсула эпифиза образована соединительной тканью мягкой мозговой оболочки. От капсулы отходят перегородки, содержащие кровеносные сосуды и сплетения симпатических нервных волокон. Эти перегородки разделяют тело железы на дольки НЕ ПОЛНОСТЬЮ. Паренхима эпифиза состоит из пинеалоцитов и интерстициальных (глиальных) клеток. Пинеалоциты содержат крупное ядро, развитую гладкую ЭПС, элементы гранулярной ЭПС, свободные рибосомы, комплекс Гольджи, много секреторных гранул, микротрубочки и микрофиламенты. Интерстициальные клетки имеют многочисленные ветвящиеся отростки, округлое плотное ядро, элементы гранулярной ЭПС и структуры цитоскелета. Вопрос 16. Гормоны эпифиза и их биологическая роль. В эпифизе синтезируются 2 гормона: Серотонин синтезируется днём, регулирует тонус ГМК, ЧСС и является медиатором некоторых нейронов ЦНС; Мелатонин синтезируется ночью, подавляет секрецию гонадотропных гормонов, препятствуя преждевременному половому созреванию. С возрастом в эпифизе накапливаются конкреции – эпифизарный песок (соли органических кислот). Вопрос 17. Развитие и строение щитовидной железы. Развитие щитовидной железы. Эндокринные клетки щитовидной железы происходят из стенки глотки и нервного гребня. На 3-4 неделе развития возникает зачаток щитовидной железы в виде выпячивания крыши глотки между первой и второй парами глоточных карманов (у корня языка). Этот зачаток растёт и к 7 неделе достигает места окончательной локализации, формируя 2 доли и перешеек. Зачаток связан с глоткой при помощи полого тяжа, открывающегося на поверхности корня языка. Часть клеток тяжа может образовывать функционально активные островки эндокринной ткани щитовидной железы. В норме этот тяж дегенерирует, оставляя после себя слепое отверстие языка (foramen caecum linguae). В конце третьего месяца развития плода начинается синтез йодсодержащих гормонов, появляющихся в амниотической жидкости. Из нервного гребня развиваются С-клетки (светлые клетки) щитовидной железы, синтезирующие кальцитонин. Строение щитовидной железы. В щитовидной железе различают строму и паренхиму. Строма состоит из: капсулы, сформированной из плотной волокнистой соединительной ткани. От капсулы внутрь органа отходят тяжи – септы, содержащие кровеносные и лимфатические сосуды, нервы; пространство органа заполняет рыхлая волокнистая соединительная ткань, поддерживающая элементы паренхимы. Она содержит капилляры фенестрированного типа, лимфатические сосуды и нервы. Паренхима щитовидной железы представлена совокупностью синтезирующих и секретирующих тиреоидные гормоны клеток и С-клеток, синтезирующих кальцитонин. Структурно-функциональной единицей щитовидной железы является ФОЛЛИКУЛ - однослойный пузырёк, образованный тиреоцитами, высота которых прямопропорциональна функциональной активности железы (в активной – цилиндрическая форма, в угнетённой – кубические или плоские). Полость фолликулов заполнена коллоидом. С-клетки (си-клетки) расположены одиночно между фолликулами. Вопрос 18. Фолликулярные клетки щитовидной железы. Образование йодсодержащих гормонов, регуляция образования. Эффекты гормонов. Фолликулярные клетки щитовидной железы (тиреоциты) образуют стенку фолликула и синтезируют тироглобулин, богатый остатками аминокислоты тирозина. После синтеза через апикальную поверхность тиреоциты секретируют тиреоглобулин в полость фолликула, где он, смешиваясь с водой, образует вязкую массу – коллоид. По учебнику. Фолликулярные клетки захватывают из кровеносного русла (щитовидная железа богато васкуляризирована) большое количество йода. Йод в тиреоцитах подвергается превращениям под действием тиропероксидазы. Этот йод так же секретируется в полость фолликула. В полости фолликула, йод связывается с тирозиновыми остатками в составе тиреоглобулина. Т. е. происходит йодирование или созревание только что синтезированного незрелого тиреоглобулина. Созревание происходит примерно 2 суток. Зрелый тиреоглобулин поглощается тиреоцитами по механизму фагоцитаза. В цитоплазме тиреоцитов капельки с тиреоглобулином сливаются с лизосомами, под действием ферментов тиреоглобулин подвергается протеолизу. Йодированные тирозиновые остатки вычленяются их тиреоглобулина и попарно коденсируются с образование трийодтиронина (Т3) и тетрайодтиронина (Т4, тироксин). Готовые гормоны секретируются в кровь. Оставшиеся аминокислоты используются для повторного синтеза тиреоглобулина. Часть Т3 образуется в печени в результате дейодинирования Т4. Т3 в 4 раза активнее Т4. По лекции Валлиулина В. В. Тиреоглобулин захватывается из полости фолликула. Используя лизосомальный аппарат в тиреоцитах происходит вычленение тирозиновых остатков из тиреоглобулина. После этого тирозиновые остатки попарно конденсируются и начинается процесс йодирования с последовательным образованием моно-, ди-, три- и тетра-йодтиронина. Т1, Т2 не активны. Т3 активнее Т4 в 40 (в сорок!) раз. Эффект Т3 и Т4. Ускоряют катаболизм, необходимы для нормального развития ЦНС как в пренатальный так и в постнатальный период. Синтез белков увеличивается в результате транскрипции в клетках мишенях. Возрастает потребление кислорода в результате увеличения активности Na+, K+ - АТФазы. Синтез управляется ТТГ. Так же синтез невозможен при недостатке йода. Вопрос 19. С-клетки щитовидной железы. Локализация, строение, гормоны, их действие. С-клетки (читается как «Си-клетки»), еще их называют «светлые клетки» или «парафолликулярные клетки». С-клетки в щитовидной железе расположены одиночно, примыкая к фолликулам или располагаясь между ними. С-клетки крупней тироцитов и имеют светлую цитоплазму. Имеют морфологию типичную для клеток, синтезирующих белок на экспорт. Комплекс Гольджи, гр ЭПС, секреторные гранулы, митохондрии. Гормоны С-клеток – кальцитонин и катакальцин. Этот гормон понижает концентрацию кальция в крови за счет активации деятельности остеобластов и угнетения функции остеокластов. Катакальцин оказывает тот же эффект, что и кальцитонин. Тропным фактором для этих гормонов является концентрация кальция в крови. Регуляция осуществляется по принципу прямой связи. Вопрос 20. Развитие и строение паращитовидной железы, клеточные типы, гормоны и их эффекты. Развитие. Нижние паращитовидные железы развиваются из энтодермы третьей пары глоточных карманов, а верхние – из энтодермы четвёртой пары. Строение. Четыре паращитовидные железы расположены на задней поверхности щитовидной железы ПОД ЕЁ КАПСУЛОЙ. Каждая паращитовидная железа имеет также собственную капсулу, от которой внутрь органа отходят септы, содержащие кровеносные сосуды. Паренхима железы образована тяжами и островками паратиреоцитов, между которыми залегают многочисленные капилляры, и содержит ГЛАВНЫЕ и ОКСИФИЛЬНЫЕ клетки, а также жировые клетки. Клеточный состав. Главные клетки паращитовидной железы имеют базофильную цитоплазму с развитой гранулярной ЭПС, комплексом Гольджи, мелкими митохондриями и секреторными гранулами. Секреторные гранулы этих клеток содержат паратиреоидный гормон (ПТГ, парат-гормон, паратиреокрин). Оксифильные клетки равномерно распределены в паренхиме паращитовидной железы или образуют скопления. Они содержат крупные митохондрии, слабо выраженный комплекс Гольджи и гранулярную ЭПС. Гормоны и их эффекты. Паратиреоидный гормон поддерживает гомеостаз кальция: увеличивает содержание кальция в сыворотке, усиливая его вымывание из костей, за счет активации деятельности остеокластов и ингибирования деятельности остеобластов и канальцевую реабсорбцию в почках; стимулирует образование кальцитриола в почках, который усиливает всасывание кальция в кишечнике; уменьшает реабсорбцию фосфатов в канальцах почки и усиливает их вымывание из костей.
Вопрос 21. Развитие надпочечника. На 6 неделе внутриутробного развития мезодермальные клетки целомического эпителия образуют скопление на краниальном конце мезонефроса, после чего формируется сосудистый полюс – место проникновения в центральную часть железы мигрирующих из нервного гребня будущих хромафинных клеток мозгового вещества. На 8 неделе мезодермальные клетки начинают интенсивно размножаться, и формируется две зоны коры: Фетальная (эмбриональная) кора располагается на границе с мозговым веществом. Клетки этой зоны у плода крупные, с ацидофильной цитоплазмой и большим ядром. К 30 неделе объём этой зоны увеличивается вдвое, но незадолго до рождения начинается дегенерация фетальной зоны коры надпочечника, и к концу первого года жизни она полностью исчезает. Функция: здесь синтезируются кортизол и дегидроэпиандростерон, который преобразуется в печени в 16а-произаодные, из которого в плаценте образуется большая часть эстрогенов материнского организма (эстриол, эстрадиол, эстрон). Дефинитивная кора располагается снаружи. Клетки этой зоны мелкие, с базофильной цитоплазмой и плотным ядром. К 30 неделе объём этой зоны значительно увеличивается, а в течение первого года жизни в дефинитивной коре различают клубочковую, пучковую и сетчатую зоны. Полная дифференцировка корковой части надпочечника завершается к третьему году жизни, но кора достигает окончательных размеров только к 20 годам жизни, особенно мощно увеличиваясь в объёме при половом созревании. Мозговая часть надпочечника к 30 неделе увеличивается в объёме в 4 раза, и вплоть до полового созревания возрастает число хромафинных клеток. Хромафинные клетки являются производными нервного гребня. Функция: хромафинные клетки плода сильно чувствительны к изменениям гомеостаза, отвечая на эти изменения выбросом катехоламинов. Вопрос 22. Строение коркового вещества надпочечника. Надпочечник окружён капсулой из плотной волокнистой соединительной ткани, от которой в толщу железы отходят соединительнотканные перегородки. Строма надпочечника состоит из рыхлой волокнистой соединительной ткани, поддерживающей эндокриноциты и содержащей большое количество кровеносных капилляров фенестрированного типа. В связи с тем, что эпителиальные тяжи паренхимы надпочечника имеют различную ориентацию в разных участках коркового слоя, которые также отличаются характером синтеза гормонов, в коре надпочечника выделяют 3 зоны: Клубочковая зона (15% всей коры) – пучки эндокриноцитов подворачиваются под капсулу и на срезе имеют вид клубочка. Эндокринные клетки данной зоны синтезируют минералокортикоиды (альдостерон, секретирующийся ТОЛЬКО В КЛУБОЧКОВОЙ ЗОНЕ); Пучковая зона (75% всей коры) – тяжи эндокринных клеток и находящиеся между ними кровеносные капилляры расположены параллельно друг другу, в виде пучков. Спонгиоциты (эндокриноциты данной зоны) синтезируют глюкокортикоиды (кортизол и кортизон), а также андрогены; Сетчатая зона (10% всей коры) – тяжи эндокриноцитов переплетаются, образуя подобие сети. Здесь синтезируются глюкокортикоиды и стероидные гормоны типа андрогенов. На границе клубочковой и пучковой зон расположены судано-фобные клетки, которые являются, камбиальным запасом надпочечника. Вопрос 23. Гормоны клубочковой зоны коры надпочечника, их эффекты, регуляция образования. В клубочковой зоне коры надпочечников происходит синтез минералокортикоидов (альдостерона и его производных). Функция минералокортикоидов – усиление обратного всасывания натрия и воды, после чего повышается артериальное давление. Минералокортикоиды поддерживают баланс электролитов жидкостей организма, который осуществляется посредством влияния на реабсорбцию ионов в почечных канальцах: альдостерон увеличивает реабсорбцию ионов натрия, хлора, бикарбоната, а также почечную экскрецию ионов калия и водорода. Регуляция образования: главный регулятор синтеза и секреции альдостерона - ангиотензин II (компонент системы [ренин-ангиотензин]) – стимулирует выброс альдостерона; натриуретические факторы ингибируют синтез альдостерона; гиперкалиемия стимулирует секрецию альдостерона, гипокалиемия – её тормозит; травмы и стрессовые состояния увеличивают секрецию альдостерона. Вопрос 24. Гормоны пучковой зоны коры надпочечника, их эффекты, регуляция образования. В пучковой зоне коры надпочечника синтезируются глюкокортикоиды (кортизол – 80%; кортизон, кортикостерон и др. – 20%). Функции глюкокортикоидов: стимулируют образование глюкозы в печени путем увеличения скорости глюконеогенеза и стимуляции освобождения аминокислот в мышцах; выступают как иммунодепрессанты (в высоких дозах); имеют выраженный противовоспалительный эффект (в малых дозах); при длительном их применении ингибируют синтетическую активность фибро- и остеобластов, в результате чего развиваются истончение кожи и остеопороз; длительное их применение поддерживает катаболизм мышц, что приводит к их атрофии и мышечной слабости; могут уменьшить отёк слизистой оболочки воздухоносных путей. Вопрос 25. Гормоны сетчатой зоны коры надпочечника, их эффекты, регуляция образования. В сетчатой зоне коры надпочечников синтезируются глюкокортикойды (описаны выше) и андрогены. Андрогены и у мужчин и у женщин в надпочечниках синтезируются одни и те же: тестостерон (большая часть), прогестерон и эстрогены. Тестостерон. Эффект: Необходим для половой дифференцировки, полового созревания, поддержания сперматогенеза. Кроме надпочечников синтезируется в клетках Лейдига яичек (большая часть). Синтез и секреция тестостерона стимулируется лютеинизирующим гормоном (ЛГ). Эстрогены. Определяют течение овариальноменструального цикла у женщин. Образуется в фолликулярных клетках яичника из тестостерона (большая часть). Прогестерон. Синтезируется клетками желтого тела и клетками плаценты при беременности. Эффект: Обеспечивает формирование функционального эндометрия в лютеиновую стадию менструального цикла; уменьшает порог возбудимости ГМК миометрия; поддерживает тонус ГМК шейки матки. Стимулируют синтез прогестерона лютропин и хорионический гонадотропин человека (ХГЧ), синтезируемый в плаценте в период беременности. Вопрос 26. Кровоснабжение и иннервация надпочечника. Кровоснабжение надпочечника обильно и осуществляется: верхней надпочечниковой артерией, являющейся ветвью нижней диафрагмальной; средней надпочечниковой артерией, отходящей от аорты; нижней надпочечниковой артерией, являющейся ветвью почечной артерии. Венозный отток коры надпочечника происходит через синусоиды мозговой части, что объясняет сочетанное вовлечение органа в развитие стрессовых ситуаций, т.к. глюкокортикоиды коры стимулируют секрецию адреналина из хромафинных клеток. Иннервация. Мозговая часть надпочечника имеет множество преганглионарных нервных волокон симпатического отдела нервной системы, а хромафинные клетки расцениваются как постганглионарное звено двигательной вегетативной иннервации. Т. е. вегетативные нервные волокна, иннервирующие надпочечник не переключаются на постганглионарных нейронах. Вопрос 27. Строение мозгового вещества надпочечника. Строма мозгового вещества надпочечника представлена каркасом из рыхлой волокнистой соединительной ткани, в которой расположены многочисленные сосудистые полости – венозные синусы. Среди трабекул расположены выполняющие эндокринную функцию хромаффинные клетки. Хромаффинные клетки крупные, овальные и объединены в гроздья и неправильной форм тяжи, располагающиеся вокруг кровеносных сосудов. Эти клетки содержат многочисленные митохондрии, выраженный комплекс Гольджи, гр ЭПС, многочисленные электроноплотные кранулы содержащие продукты синтеза (адреналин, норадреналин, эндорфин, энкефалины). Вопрос 28. Гормоны мозгового вещества надпочечника, их биологическая роль. В мозговом веществе надпочечника вырабатываются: Катехоламины: адреналин, норадреналин, дофамин. Они регулируют ЧСС, тонус ГМК, выработку глюкокортикоидов;
Опиоидные пептиды: эндорфины и энкефалины, являющиеся естественными анальгетиками, уменьшающими болевую чувствительность. Вопрос 29. Организация и клеточный состав панкреатического островка (Лангерганса). Панкреатические островки (островки Лангерганса) представляют собой эндокринную часть поджелудочной железы. Разбросаны по всей железе в виде округлых компактных скоплений эндокринных клеток (инсулоцитов) между ацинусами (экзокринная часть поджелудочной железы). Общее количество островков у взрослых составляет 0,5-2 млн. в хвосте железы островков больше. Островки Лангерханса состоят из нескольких сотен или тысяч инсулоцитов, между которыми располагаются ретикулярные волокна, фенестрированые кровеносные капилляры и нервные волокна. Инсулоциты характеризуются овальной или полигональной формой, развитым синтетическим аппаратом, образованием и накоплением в цитоплазме окруженных мембраной секреторных гранул различных размеров, формы и плотности, содержащих полипептидные гормоны. Клеточные типы смотри ниже! Вопрос 30. Клетки панкреатического островка (Лангерганса), их гормоны, эффекты гормонов. Эндокринная часть – выработка гормонов. Включает пакреатические островки, состоящие из инсулиноцитов: α-клетки - в гранулах глюкагон, под его влияние гликоген распадается на простые сахара, β-клетки - выделение инсулина, стимулирует усвоение клетками простых сахаров, синтез гликогена из простых сахаров и депонирование в печени (при избытки снижается уровень сахара в крови), D-клетки - соматостатины, задерживают выделение инсулина и глюкагона, ингибирует синтез ферментов ацинозными клетками, D1-клетки - вазоактивный интерстинальный пептид, снижающий АД и стимулирующий секрецию ферментов и гормонов пожделудочной железы, PP-клетки – панкреатический полипептид, стимулирующий выделение желудочного и панкреатического сока. Вопрос 31. Строение жёлтого тела яичника, его гормоны, их мишени и эффекты гормонов. Строение. Внутри желтого тела находится соединительнотканный рубец, окруженный лютеиновыми клетками, между которыми расположены капилляры синусоидного типа. Функции. Желтое тело функционирует в лютеиновую фазу овариального цикла. Под действием ЛГ лютеиновые клетки синтезируют и секретируют прогестерон. Также лютеиновые клетки секретируют эстрогены (но прогестерона больше: прогестерона 2/3, а эстрогенов 1/3). Менструальное жёлтое тело функционирует до завершения цикла (имплантации нет). Уровень прогестерона достигает пика через 8–9 дней после овуляции, что приблизительно соответствует времени имплантации (если происходит оплодотворение яйцеклетки). Жёлтое тело беременности. ХГТ (хорионический гонадотропин), секретируемый клетками трофобласта (плаценты), через рецепторы ЛГ лютеиновых клеток стимулирует рост жёлтого тела, достигающего в диаметре 5 см, и увеличивает продукцию половых гормонов. Высокий уровень прогестерона и эстрогенов сохраняет развивающуюся беременность. Также в клетках этого желтого тела синтезируется релаксин. Желтое тело беременности наиболее активно в период первого триместра беременности. Функции гормонов. Релаксин – в течение беременности оказывает расслабляющий эффект на миометрий, перед родами приводит к расширению маточного зева и уменьшению плотности лонного сочленения. Эстрогены. Определяют течение овариальноменструального цикла у женщин. Прогестерон. Обеспечивает формирование функционального эндометрия в лютеиновую стадию менструального цикла; уменьшает порог возбудимости ГМК миометрия; поддерживает тонус ГМК шейки матки. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||