Гистология реферат. Тема 1 клетка Вопрос Химический состав, организация плазмолеммы. Функции плазмолеммы
 Скачать 3.29 Mb. Скачать 3.29 Mb.
|

|
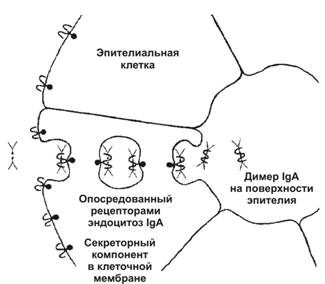
Тема 16: «Иммунная защита» Вопрос 1. Антитело как белок семейства иммуноглобулинов. Структура мономера молекулы иммуноглобулина. Основные классы иммуноглобулинов, особенности их строения и функции. Антитело – гликопротеин, относящийся к классу иммуноглобулинов, молекула которого имеет уникальный участок связывания эпитопа (антигенной детерминанты). Антитела синтезируются в плазматических клетках в ходе гуморального иммунного ответа. Молекула иммуноглобулина состоит из двух лёгких цепей и двух тяжёлых цепей, в которых различают: вариабельную область (вариабельные области легкой и тяжёлой цепи образуют антиген-связывающий центр – Fab-фрагмент, с которым взаимодействует эпитоп иммуногена); константную область, имеющую Fc-фрагмент, который связывается с клетками-эффекторами (макрофаги, тучные клетки и др.), несущими на своей поверхности рецепторы Fc-фрагмента. 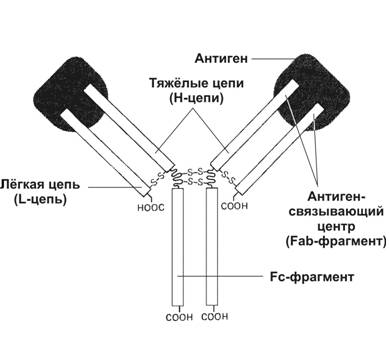 В зависимости от структуры тяжёлых цепей, принято выделять 5 классов иммуноглобулинов: IgА – основной класс антител в слюне, слезе, молоке; выделяется на поверхность слизистых, где взаимодействует с антигеном. Молекулы IgA переносятся через эпителиальную клетку во внешнюю среду.
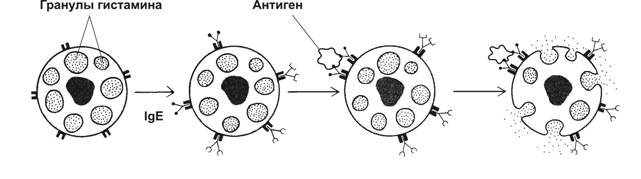
IgD – биологическая роль не установлена, появляется на поверхности развивающихся Т-лимфоцитов, содержится в сыворотке. IgE специфически взаимодействует с тучными клетками и базофильными лейкоцитами, дегрануляция которых вызывает расширение просвета венул и увеличение проницаемости их стенок. Он защищает организм от паразитов.
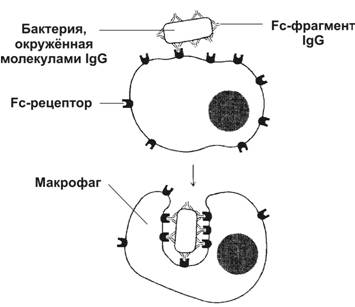
IgG – преобладающий класс антител, его много при вторичном иммунном ответе, он защищает ткани от бактерий, вирусов и токсинов.
NB! Только IgG способны проходить через плацентарный барьер, обеспечивая передачу пассивного иммунитета от матери к плоду. Механизм:
IgM продуцируется развивающимися В-клетками при первичном попадании антигена в организм. При этом он является рецептором антигенов, встраиваясь в плазматическую мембрану В-лимфоцита. Вопрос 2. Молекулы главного комплекса гистосовместимости (MHC). Функциональное значение МНС-I при клеточном иммунитете. Функциональное значение MHC-II при гуморальном ответе. Молекулы MHC-I и MHC-II – гликопротеины плазматической мембраны, экспрессированные на всех антиген-представляющих клетках. Они являются мишенями иммунного ответа при отторжении трансплантата. Спектр молекул MHC строго индивидуален для каждого, но в пределах одного организма он абсолютно идентичен для всех соматических клеток. Таким образом, главный комплекс гистосовместимости определяет биологическую индивидуальность организма. Молекула МНС-I представлена на мембранах ВСЕХ ЯДРОСОДЕРЖАЩИХ клеток и состоит из альфа-цепи, закрепленной в клеточной мембране. Внемембранная часть этой цепи связана с короткой цепью бета2-микроглобулина, не имеющего трансмембранного участка. Молекула МНС-II имеется только на мембранах иммунокомпетентных клеток и состоит из двух субъединиц: более длинной альфа-цепи и бета-цепи. Каждая цепь содержит небольшой фрагмент в цитоплазме, трансмембранный участок, а также часть, выступающую на поверхности клеточной мембраны. Функции МНС-I при клеточном иммунном ответе: молекулы МНС-I взаимодействуют с молекулой CD8, которая находится на мембране предшественника цитотоксического Т-лимфоцита. Функции МНС-II при гуморальном иммунном ответе: молекулы МНС-II способствуют представлению антигена Т-клеткам с помощью макрофагов молекулы МНС-II взаимодействуют с молекулой CD4, которая находится на мембране Т-хелпера, что вызывает выделение лимфокинов, которые стимулируют пролиферацию и созревание предшественников цитотоксических Т-лимфоцитов. Кроме того, МНС-II участвует во взаимодействии Т- и В-лимфоцитов. Вопрос 3. Иммунокомпетентные клетки, их общая характеристика. Т-лимфоциты состоят из функциональных подтипов CD4 и CD8. Они узнают антиген, предварительно процессированный и представленный на поверхности антигенпредставляющих клеток. Они ответственны за клеточный иммунитет, а также помогают В-лимфоцитам при гуморальном иммунном ответе реагировать на антиген. Т-хелперы (CD4) – синтезируют и секретируют цитокины - пептидные молекулы, передающие сигнал от одной клетки к другой, расположенной рядом. К цитокинам относят ИНТЕРЛЕЙКИНЫ-2, 4, 5, 6, а также гамма-ИНТЕРФЕРОН. В ходе иммунного ответа Т-хелперы узнают молекулы MHC-II. Я вляются единственной мишенью вируса СПИДа; Цитотоксические Т-лимфоциты, или Т-киллеры, или Т-эффекторы (CD-8) уничтожают инфицированные вирусом и чужеродные клетки при помощи перфорина. Перфорины – цитотоксические белки, имеющие литическую область, с помощью которой они проникают в плазматическую мембрану клетки-мишени, где соединяясь друг с другом, образуют пору, тем самым разрушая клетку-мишень. Кроме того, цитотоксические Е-лимфоциты взаимодействуют с молекулой MHC-I в плазматической мембране клетки-мишени; Т-супрессоры (CD-8) регулируют интенсивность иммунного ответа, подавляя активность Т-хелперов, предотвращают развитие аутоиммунных реакций, обеспечивают невосприимчивость матери к отцовским антигенам, представленным на клетках плода; Т-лимфоциты памяти (CD-8) обеспечивают развитие клеточного иммунитета при повторном попадании антигенов. В-лимфоциты ответственны за ГУМОРАЛЬНЫЙ ИММУННЫЙ ОТВЕТ. В их мембране имеются молекулы IgM, являющиеся рецепторами антигенов. В-лимфоциты мигрируют из красного костного мозга в тимус-независимые зоны лимфоидных органов. Продолжительность их жизни составляет менее 10 дней, если они не активируются антигенами. В-лимфоциты памяти обеспечивают развитие гуморального иммунитета при повторном попадании антигенов.
NK-клетки (5-15%) убивают ауто-, алло- и ксеногенные опухолевые клетки, а также некоторые инфицированные вирусом и бактериями клетки. Они НЕ ИМЕЮТ ПОВЕРХНОСТНЫХ ДЕТЕРМИНАНТ. В них экспрессируются дифференцировочные антигены CD2, CD56, CD16 (рецептор Fc-фрагмента). NK-клетки уничтожают клетку-мишень при помощи перфорина после установления с ней прямого контакта, а не путём фагоцитоза, т.к. не имеют ни MHC-I, ни MHC-II. Активность NK-клеток регулируется цитокинами, её усиливают интерлейкин-2 и гамма-интерферон. NK-клетки участвуют в антитело-зависимом клеточно-опосредованном цитолизе, экспрессируя на своей поверхности рецептор Fc-фрагмента IgG, с которым будет взаимодействовать Fc-фрагмент антител, связанных с клеткой-мишенью. Антиген-представляющие клетки (макрофаги, фолликулярные отростчатые клетки лимфоузлов и селезёнки, клетки Лангерганса кожи, М-клетки в лимфатических фолликулах пищеварительного тракта, дендритные эпителиальные клетки тимуса) ЗАХВАТЫВАЮТ, ПРОЦЕССИРУЮТ И ПРЕДСТАВЛЯЮТ антигены (эпитоп) на своей поверхности другим иммунокомпетентным клеткам, вырабатывают цитокины (ИЛ-1 и др.). Они секретируют простагландин Е2, угнетающий иммунный ответ, и гамма-интерферон, усиливающий фагоцитарную и цитолитическую активность макрофагов. Вопрос 4. Антигенпредставляющие клетки. Структурная характеристика, локализация в разных тканях и органах, функция. Антиген-представляющие клетки: макрофаги, фолликулярные отростчатые клетки лимфоузлов и селезёнки, клетки Лангерганса кожи, М-клетки в лимфатических фолликулах пищеварительного тракта, дендритные эпителиальные клетки тимуса, В-лимфоциты. Для антиген-представляющих клеток характерно наличие митохондрий, гранулярной ЭПС, комплекса Гольджи и лизосом. На их поверхности экспрессированы молекулы MHC-II. Функции. ЗАХВАТЫВАЮТ, ПРОЦЕССИРУЮТ И ПРЕДСТАВЛЯЮТ антигены (эпитоп) на своей поверхности другим иммунокомпетентным клеткам, вырабатывают цитокины (ИЛ-1 и др.). Они секретируют простагландин Е2, угнетающий иммунный ответ, и гамма-интерферон, усиливающий фагоцитарную и цитолитическую активность макрофагов. Вопрос 5. Типы Т-лимфоцитов. Дифференцировка (обучение) в тимусе, функция. Т-лимфоциты состоят из функциональных подтипов CD4 и CD8. Они узнают антиген, предварительно процессированный и представленный на поверхности антигенпредставляющих клеток. Они ответственны за клеточный иммунитет, а также помогают В-лимфоцитам при гуморальном иммунном ответе реагировать на антиген. Т-хелперы (CD4) – синтезируют и секретируют цитокины - пептидные молекулы, передающие сигнал от одной клетки к другой, расположенной рядом. К цитокинам относят ИНТЕРЛЕЙКИНЫ-2, 4, 5, 6, а также гамма-ИНТЕРФЕРОН. В ходе иммунного ответа Т-хелперы узнают молекулы MHC-II. Я вляются единственной мишенью вируса СПИДа; Цитотоксические Т-лимфоциты, или Т-киллеры, или Т-эффекторы (CD-8) уничтожают инфицированные вирусом и чужеродные клетки при помощи перфорина. Перфорины – цитотоксические белки, имеющие литическую область, с помощью которой они проникают в плазматическую мембрану клетки-мишени, где соединяясь друг с другом, образуют пору, тем самым разрушая клетку-мишень. Кроме того, цитотоксические Е-лимфоциты взаимодействуют с молекулой MHC-I в плазматической мембране клетки-мишени; Т-супрессоры (CD-8) регулируют интенсивность иммунного ответа, подавляя активность Т-хелперов, предотвращают развитие аутоиммунных реакций, обеспечивают невосприимчивость матери к отцовским антигенам, представленным на клетках плода; Т-лимфоциты памяти (CD-8) обеспечивают развитие клеточного иммунитета при повторном попадании антигенов. Антигеннезависимая дифференцировка (обучение) лимфоцитов в тимусе происходит в корковом веществе, а в кровоток лимфоциты запускаются в мозговом веществе тимуса. Клетка-предшественница Т-лимфоцитов поступает в тимус из костного мозга в плодном периоде. Она экспрессирует на своей поверхности дифференцировочный антиген CD7, после чего синтезирует цитоплазматическую форму CD3 (cCD3), а позже выставляет на поверхности CD1 и CD2. Таким образом, протимоциты имеют фенотип: CD7+CD1+CD2+сCD3+CD4-CD8-. Предшественники лимфоцитов, пришедшие обучаться в тимус, образуют несколько популяций: большая часть тимоцитов к обучению не способны, значительная часть способна к обучению, но в процессе обучения остаётся плохо обученной. Обе популяции уничтожаются в тимусе путём апоптоза. Только малочисленная часть тимоцитов нормально дифференцируется, выходит за пределы тимуса и обеспечивает нормальную работу иммунной системы. Когда клетка становится зрелой, молекула CD1 исчезает, а сCD3 переходит (из цитоплазматической формы) в мембранную. Далее тимоциты начинают экспрессировать CD4 и CD8, давая начало тимоцитам с фенотипом CD1+CD2+CD4+CD8+, которые способны дифференцироваться в двух направлениях: CD1-CD2+CD4+CD8- или CD1-CD2+CD4-CD8+ при наличии у обоих подтипов мембранного маркёра CD3 и рецептора Т-лимфоцита. Такие клетки покидают тимус и появляются в периферической крови и лимфоидных органах.
Вопрос 6. В-лимфоциты. Стадии развития, функции. Структура и функция плазматических клеток. Стадии развития В-лимфоцитов – их дифференцировка в Ig-продуцирующие плазматические клетки. Стволовые клетки костного мозга проходят серию этапов дифференцировки, превращаясь в зрелые В-лимфоциты:
В-лимфоциты ответственны за ГУМОРАЛЬНЫЙ ИММУННЫЙ ОТВЕТ. В их мембране имеются молекулы IgM, являющиеся рецепторами антигенов. В-лимфоциты мигрируют из красного костного мозга в тимус-независимые зоны лимфоидных органов. Продолжительность их жизни составляет менее 10 дней, если они не активируются антигенами. В-лимфоциты памяти обеспечивают развитие гуморального иммунитета при повторном попадании антигенов. Для плазматической клетки характерно наличие хорошо развитых гранулярной ЭПС и комплекса Гольджи, что свидетельствует об активном синтезе и секреции белка. Зрелые В-лимфоциты, или плазматические клетки – ЕДИНСТВЕННЫЕ клетки организма, способные к синтезу и секреции антител (Ig). Вопрос 7. NK-клетки. Образование, строение, функционирование. NK-клетки составляют 5-15% всех циркулирующих в крови лимфоцитов. Образование NK-клеток. NK-клетки не проходят дифференцировку в тимусе, они выходят из костного мозга в кровь и затем мигрируют в ткани, где осуществляют врожденную иммунную защиту, которая особенно важна для защиты организма от вирусов и опухолей. Строение NK-клеток. NK-клетки (фенотип: CD3-CD16+CD56+CD94+) – это большие зернистые лимфоциты, основная часть обильной цитоплазмы которых содержит несколько митохондрий, свободные рибосомы с отдельными элементами шероховатой ЭПС, аппарат Гольджи и характерные цитолитические гранулы с перфорином. Они лишены характерных для Т- и В-лимфоцитов поверхностных детерминант. Одной из характеристик NK-клеток является наличие Fc-рецептора. Функции NK-клеток. Основная обязанность киллерных клеток - выявлять и уничтожать собственные клетки организма, в которых что-то нарушилось: они убивают опухолевые клетки и клетки, зараженные вирусами или другими чужеродными агентами. NK-клетки уничтожают клетку-мишень при помощи перфорина после установле-ния с ней прямого контакта, а не путём фагоцитоза, т.к. не имеют ни MHC-I, ни MHC-II. Активность NK-клеток регулируется цитокинами, её усиливают интерлейкин-2 и гамма-интерферон. NK-клетки участвуют в антитело-зависимом клеточно-опосредованном цитолизе, экспрессируя на своей поверхности рецептор Fc-фрагмента IgG, с которым будет взаимо-действовать Fc-фрагмент антител, связанных с клеткой-мишенью. Вопрос 8. Антигентранспортирующие клетки (М-клетки) пищеварительного тракта: структура, функционирование. Одним из механизмов поступления антигенов из просвета кишки является их транспортировка через М-клетки, которые являются разновидностью энтероцитов, расположены над Пейеровыми бляшками. М-клетки не имеют микроворсинок, но имеют микроскладочки, с помощью которых они захватывают антигены из просвета, после чего путем эндоцитоза они транспортируют антиген через клетку. В процессе транспортировки происходит обнажение антигенных структур вещества на базолатеральной мембране (базальная мембрана и мембрана боковых отделов клетки) происходит стимуляция дендритных клеток в верхней части Пейеровой бляшки антиген представляется Т-лимфоцитам. Антигены, представленные Т-хелперам и макрофагам, распознаются и, в случае наличия на поверхности этих клеток соответствующих антигену рецепторов, недифференцированные T-хелперы трансформируются в 2 типа: Трансформация в T-хелперы I типа сопровождается выработкой цитокинов, активацией фагоцитоза, миграцией нейтрофилов, усилением окислительных реакций, синтезом IgА. Эти реакции направлены на уничтожение антигена. Дифференцировка во II тип T-хелперов способствует выработке противовоспалительных цитокинов, выработку IgG, а также способствует образованию IgЕ. Вопрос 9. Взаимодействия клеток при гуморальном и клеточном иммунном ответе. Гуморальный иммунный ответ: макрофаг + T-хелпер + В-лимфоцит
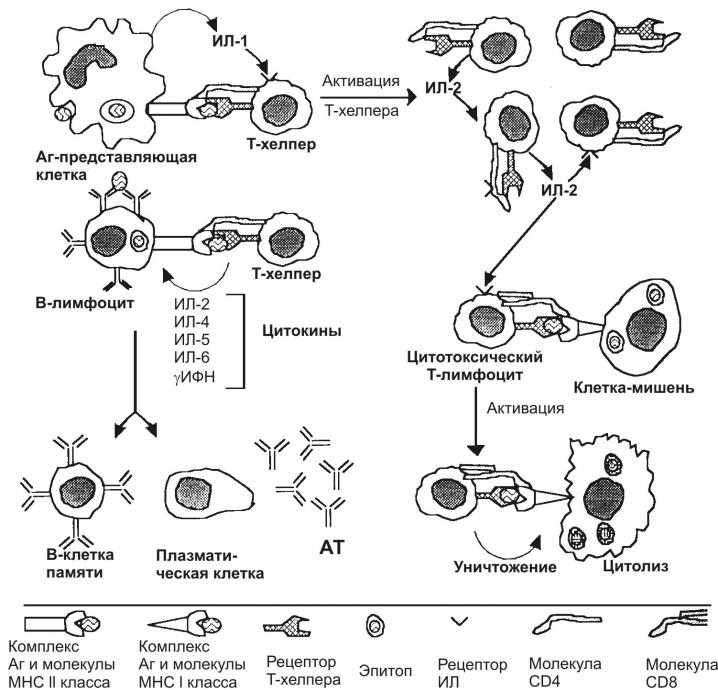 Клеточный иммунный ответ: цитотоксический Т-лимфоцит
Вопрос 10. Вилочковая железа (тимус): развитие, строение, функции. Гемато-тимический барьер: структура, функции. Гормоны тимуса. Развитие. Эпителий бранхиогенной группы желёз, к которой, помимо щитовидной и паращитовидной, относят также тимус, развивается из энтодермы глоточных карманов: вилочковая железа развивается из ТРЕТЬЕЙ и ЧЕТВЁРТОЙ пары ГЛОТОЧНЫХ КАРМАНОВ. Закладка растёт в каудально-вентральном направлении, сохраняя контакт с глоткой, после чего отделяется от нее и смещается каудально и медиально с последующим слиянием по средней линии с закладкой другой стороны. Вскоре в зачатке тимуса появляются и начинают интенсивно размножаться лимфоидные клетки, мигрирующие из костного мозга. Большинство эпителиальных клеток тимуса происходит из эпителиальных (энтодермальных) стволовых клеток, но также они могут образовываться и из эктодермы. Строение. Капсула тимуса и отходящие от нее внутрь органа септы построены из плотной волокнистой соединительной ткани. Объём тимуса заполнен эпителиальным каркасом, в котором располагаются тимоциты. В дольке зрелого тимуса различают корковый и мозговой слои: Корковый слой содержит делящиеся клетки – лимфобласты (предшественники Т-лимфоцитов), которые взаимодействуют с дендритными эпителиальными клетками. Дендритные эпителиальные клетки имеют длинные отростки, соединяющиеся между собой при помощи десмосом, а в их цитоплазме имеются гранулы, содержащие гормоны тимуса: тимозины и тимопоэтин. Кроме того, эти клетки экспрессируют большое количество молекул МНС-II. Функционально важна внутренняя часть коркового слоя, которая содержит потомки лимфобластов, неделящиеся малые тимоциты и дендритные эпителиальные клетки. По мере созревания, что происходит именно во внутренней части коркового слоя тимуса, протимоциты утрачивают дифференцировочный антиген CD1, но приобретают CD3, CD4 и CD8. Дальнейшая их дифференцировка протекает в мозговой части дольки тимуса. Мозговой слой принимает тимоциты, поступающие из коркового слоя тимуса, где они дифференцируются в CD4+ и CD8+ лимфоциты. Зрелые Т-клетки (в процессе обучения их остаётся 3-5% от общего количества поступившего в тимус из красного костного мозга клеток-предшественниц Т-лимфоцитов) выходят из мозгового слоя по венулам и выносящим лимфатическим сосудам. Остальные клетки фагоцитируются макрофагами, находящимися в мозговом слое тимуса.
Функции тимуса: уничтожение лимфоцитов, способных узнавать антиген собственного организма. Молекулы рецепторов в мембране тимоцита взаимодействуют с комплексом МНС-аутоантиген в мембране эпителиальной клетки. Клоны тех тимоцитов, рецепторы которых узнают комплекс [МНС-аутоантиген], уничтожаются. выработка гуморальных факторов иммунной системы: тимозин и тимопоэтин. Тимозины способствуют дифференцировке Т-лимфоцитов и появлению специфических рецепторов в их мембране; они стимулируют выработку лимфокинов и продукцию иммуноглобулинов. Тимопоэтин является стимулятором дифференцировки предшественников Т-лимфоцитов, а также влияет на дифференцировку Т-лимфоцитов. Вопрос 11. Селезенка: развитие, строение, функции. Кровеносные сосуды селезенки. Структурно-функциональная организация красной и белой пульпы. Гормоны селезенки. Развитие. Закладка селезенки происходит на 5-й неделе эмбриогенеза образованием плотного скопления мезенхимы. Последняя дифференцируется в ретикулярную ткань, прорастает кровеносными сосудами, заселяется стволовыми кроветворными клетками. На 5-м месяце эмбриогенеза в селезенке отмечаются процессы миелопоэза, которые к моменту рождения сменяются лимфоцитопоэзом. 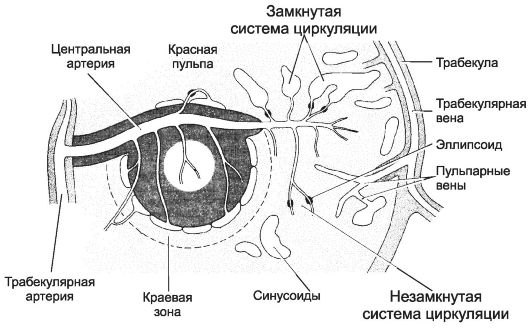 Строение. Селезёнка снаружи покрыта соединительнотканной капсулой, содержащей ГМК и много эластина. От капсулы внутрь органа отходят трабекулы, содержащие артерию и растянутые кровью трабекулярные вены. Между трабекулами расположена паренхима органа, в которой выделяют: красную пульпу, окружающую лимфатические фолликулы. Здесь преобладают эритроциты, есть много макрофагов, уничтожающих отжитые эритроциты; эллипсоиды и венозные синусы растянуты заполняющей их кровью. Ткань фолликулов располагается вокруг центральной артерии. Лимфатические фолликулы – тимус-независимая зона. В нём выделяют 2 зоны: центр размножения – центральная часть фолликула, где присутствуют макрофаги, фолликулярные отростчатые клетки и В-лимфоциты; краевая зона – граница между фолликулом и красной пульпой, где присутствуют активно фагоцитирующие макрофаги. Во внутренней части этой зоны расположены синусы, куда поступает кровь из артериальных сосудов фолликула. Кровь вступает в контакт с паренхимой органа, из нее в ткань выходят Т- и В-лимфоциты, распределяющиеся по специфическим для каждого клеточного типа зонам селезёнки. белую пульпу – совокупность лимфоидной ткани селезенки, представленной скоплением Т-лимфоцитов вокруг артерий, выходящих из трабекул (тимус-зависимая зона). Функции селезёнки: продукция иммуноглобулинов, необходимых для быстрого и эффективного удаления бактерий из кровотока. Селезёнка участвует в удалении плохо опсонизированных бактерий (хорошо опсонизированные бактерии из кровотока удаляются в печени); фагоцитоз повреждённых и старых эритроцитов; гуморальная функция: селезёнка – место образования гуморальных факторов, влияющих на систему мононуклеарных фагоцитов. Гормоны селезёнки: Тафтсин стимулирует активность фагоцитов; Спленин – функциональный аналог тимопоэтина, который является стимулятором дифференцировки предшественников Т-лимфоцитов, а также влияет на дифференцировку Т-лимфоцитов. Кровообращение в селезёнке:
Вопрос 12. Лимфатический узел: развитие, строение, функции. Структуры коркового и мозгового вещества, паракортикальная зона, синусы. Развитие. Зачатки лимфатических узлов появляются в конце 2-го - начале 3-го месяца эмбриогенеза в виде скоплений мезенхимы по ходу лимфатических сосудов. Вскоре из мезенхимы образуется ретикулярная ткань, составляющая строму органа. К концу 4-го месяца в закладки узлов вселяются лимфоциты и формируются скопления — первичные узелки без центра размножения. Одновременно появляется подразделение органа на корковое и мозговое вещество. Строение. Лимфатический узел снаружи покрыт соединительнотканной капсулой, от которой внутрь органа отходят трабекулы. В нём различают корковую и мозговую части, между которыми расположена тимус-зависимая паракортикальная зона, а также синусы. 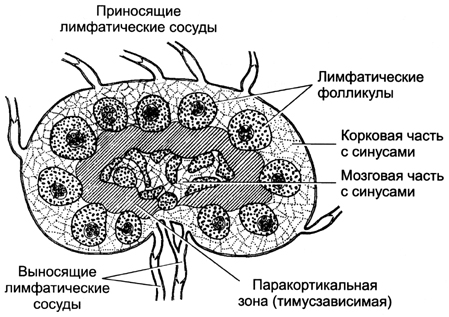 Корковое вещество. Здесь расположены скопления лимфоидной ткани в виде вторичных узелков. Это округлые образования диаметром до 1 мм. Центральную часть узелка называют центром размножения, или реактивным центром. Здесь происходит антигензависимая пролиферация В-лимфоцитов и дифференцировка их в предшественники плазматических клеток. Кроме того, в центре размножения находятся дендритные клетки костномозгового происхождения, которые на своих отростках удерживают антигены, активирующие В-лимфоциты, макрофаги моноцитарного генеза, фагоцитирующие погибающие апоптозом аутоиммунные В-лимфоциты, антигены и инородные частицы. По периферии вторичного узелка расположена корона полулунной формы, состоящая из малых лимфоцитов (рециркулирующих В-лимфоцитов, В-клеток памяти, незрелых плазматических клеток). На границе центра размножения короны обнаруживаются Т-лимфоциты (хелперы), которые способствуют развитию В-лимфоцитов в иммунобласты. Последние мигрируют в мозговые тяжи, отходящие от паракортикальной зоны и узелков внутрь мозгового вещества. Лимфатические узелки являются динамическими структурами. Они то образуются, то исчезают. Паракортикалъная зона лимфатического узла находится на границе между корковым и мозговым веществом. Она называется тимусзависимой зоной, или Т-зоной, так как при удалении тимуса происходит ее исчезновение. В паракортикальной зоне осуществляются бласттрансформация Т-лимфоцитов, их пролиферация и превращение в специализированные клетки иммунной системы. Здесь много дендритных клеток. Они появляются в результате миграции из покровных тканей внутриэпидермальных макрофагов. На своей поверхности они несут антигены и представляют их Т-лимфоцитам (хелперам). Кроме того, в этой зоне находятся особые венулы, выстланные эндотелиоцитами кубической формы. Через стенку этих венул происходит переход Т- и В-лимфоцитов из крови в строму лимфатического узла. Мозговое вещество лимфатических узлов является местом созревания плазматических клеток. Вместе со вторичными узелками коркового вещества мозговые тяжи составляют тимуснезависимую зону, или В-зону, лимфатических узлов. Мозговые тяжи кроме В-лимфоцитов и плазмоцитов содержат Т-лимфоциты и макрофаги. Синусы. Под капсулой расположен краевой синус, куда поступает лимфа из приносящих лимфатических сосудов. Краевой синус через промежуточные синусы переходит в синусы мозгового вещества, а из них лимфа по выносящим лимфатическим сосудам в области ворот выходит из органа. Тема 17: «Пищеварительная система» Вопрос 1. Сроки и источники развития структур пищеварительной трубки (ПВТ). 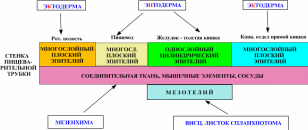 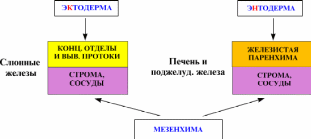 В конце 3-й недели эмбрионального развития плоский 3-х листковый зародыш человека сворачивается в трубку, т.е. формируется тело. При этом энтодерма, висцеральный листок спланхнотомов и мезенхима между ними сворачиваясь в трубку образуют I кишку – это замкнутая в краниальном и каудальном конце полая трубка, выстланная изнутри энтодермой, снаружи – висцеральным листком спланхнотомов, слоем мезенхимы между ними. В переднем отделе зародыша эктодерма впячиваясь навстречу краниальному слепому концу I кишки образует I ротовую бухту, в каудальном конце зародыша эктодерма впячиваясь навстречу другому слепому концу I кишки образует анальную бухту. Просвет I кишки от полостей этих бухт отграничен соответственно глоточной и анальной мембраной. Энтодерма переднего отдела замкнутой I кишки состоит из клеточного материала бывшей прехордальной пластинки эпибласта, остальные участки энтодермы I кишки – материал гипобласта. В заднем отделе I кишки образуется слепое выпячивание – формируется аллантоис, являющийся рудиментарным провизорным органом эмбриона человека. Глоточная и анальная мембраны в последующем прорываются и ПВТ становится проточной. Граница между эпителиями, развивающегося из материала прехордальной пластинки и из материала гипобласта четко прослеживается и соответствует линии перехода многослойного плоского неороговевающего эпителия пищевода в эпителий желудка. Из эктодермы ротовой бухты образуется эпителий предверья ротовой полости, из энтодермы переднего отдела I кишки (материал прехордальной пластинки) – эпителий ротовой полости и его производные, эпителий глотки и пищевода, эпителий дыхательной системы (трахеи, бронхиального дерева и респираторного отдела дыхательной системы); из остальной части энтодермы (материал гипобласта) образуется эпителий и железы желудка и кишечника, эпителий печени и поджелудочной железы; из эктодермы анальной бухты образуется многослойный плоский неороговевающий эпителий и эпителий желез анального отдела прямой кишки. Из мезенхимы I кишки образуется рыхлая волокнистая сдт собственной пластинки слизистой, подслизистой основы, адвинтициальной оболочки и прослойки рыхлой сдт мышечной оболочки, а также гладкая мышечная ткань (мышечная пластинка слизистой и мышечная оболочка). Из висцерального листка спланхнотомов I кишки образуется серозный (брюшинный) покров желудка, кишечника, печени и частично поджелудочной железы. Печень и поджелудочная железа закладываются как выпячивание стенки I кишки, т. е. тоже из энтодермы, мезенхимы и висцерального листка спланхнотомов. Из энтодермы образуются гепатоциты, эпителий желчевыводящих путей и желчного пузыря, панкретоциты и эпителий выводных путей поджелудочной железы, клетки островков Лангерганса; из мезенхимы образуются сдт элементы и гладкая мышечная ткань, а из висцерального листка спланхнотомов – брюшинный покров этих органов. Энтодерма аллантоиса участвует при развитии переходного эпителия мочевого пузыря. Вопрос 2. Общий план строения пищеварительной трубки. Стенка пищеварительной трубки состоит из слизистой, подслизистой, мышечной и наружной (серозная или адвентициальная) оболочек (рис. 12-18). 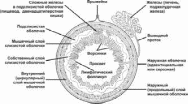 Стенка пищеварительной трубки состоит из 4-х оболочек: слизистая, подслизистая, мышечная и серозная (или адвентициальная). Слизистая оболочка содержит лимфатические фолликулы и может включать простые экзокринные железы (например, в желудке). Подслизистая оболочка некоторых отделов пищеварительного тракта (пищевод, двенадцатиперстная кишка) имеет сложные железы. Выводные протоки всех экзокринных желёз пищеварительного тракта открываются на поверхности слизистой оболочки. Через стенку пищеварительной трубки проходят и открываются на поверхности слизистой оболочки выводные протоки больших желёз пищеварительной системы — печени и поджелудочной железы. Слизистая оболочка Слизистая оболочка состоит из трёх слоёв: эпителия, собственного слоя и мышечного слоя. В пищеварительной трубке присутствуют слизистые оболочки двух типов: кожного и кишечного (табл. 12-2). Это подразделение основано на характере эпителия слизистой оболочки: слизистая кожного типа имеет многослойный плоский эпителий, слизистая кишечного типа — однослойный. Слизистая оболочка кишечного типа — часть системы иммунной защиты организма и может быть отнесена к органам лимфоидной системы. · Эпителий. Слизистая оболочка кожного типа содержит многослойный плоский эпителий, слизистая оболочка кишечного типа — однослойный цилиндрический. · Cобственный слой состоит из рыхлой волокнистой соединительной ткани. Здесь встречаются ГМК, лимфоциты, плазматические клетки, фибробласты и тучные клетки. · Мышечный слой построен из ГМК. В слизистой оболочке кожного типа мышечный слой присутствует только в пищеводе. Мышечный слой обеспечивает изменения рельефа слизистой оболочки. В нём различают два подслоя. В одном из них все ГМК ориентированы циркулярно, а в другом подслое — продольно. Таблица 12-2. Типы слизистых оболочек
Подслизистая оболочка Подслизистая оболочка находится между слизистой и мышечной оболочками, представлена рыхлой волокнистой соединительной тканью и содержит сплетение кровеносных сосудов и подслизистое (майсснеровское) нервное сплетение. Оба сплетения необходимы для обеспечения жизнеспособности и выполнения функций слизистой оболочки. Подслизистая оболочка реализует локальные смещения слизистой оболочки относительно мышечной, отсутствует в дёснах, твёрдом нёбе и на дорсальной (верхней) поверхности языка. Мышечная оболочка На всём протяжении пищеварительного тракта, начиная от границы между верхней и средней третью пищевода, мышечная оболочка построена из ГМК, которые образуют два слоя: циркулярный и продольный. Сокращение ГМК продольного слоя укорачивает длину кишечной трубки. ГМК циркулярного слоя развивают сфинктероподобные сокращения. Между этими слоями расположено ауэрбаховское нервное сплетение. ГМК участвуют как в физиологической, так и в репаративной регенерации, способны к синтезу коллагенов, эластина и других компонентов межклеточного вещества. Наружная оболочка Если рассматриваемая часть пищеварительного тракта обращена в брюшную полость, то наружная оболочка — серозная. T. serosa везде устроена однотипно: со стороны брюшной полости (плевральной полости, полости сердечной сумки) — пласт однослойного плоского эпителия (мезотелия); под его базальной мембраной расположена пластинка рыхлой волокнистой соединительной ткани. Мезотелий обладает хорошими транспортными свойствами (например, для всасывания из полости избытка жидкости). Серозная оболочка обеспечивает беспрепятственное скольжение находящихся в полости органов относительно друг друга. Вопрос 3. Строение слизистой оболочки и ее особенности в различных отделах пищеварительной трубки. Свое название она получила в связи с тем, что поверхность ее постоянно увлажняется выделяемой железами слизью. Эта оболочка состоит, как правило, из трех пластинок: эпителиальной пластинки (эпителия), собственной пластинки слизистой (lamina propria mucosae) и мышечной пластинки слизистой (lamina muscularis mucosae). Эпителий в переднем и заднем отделах пищеварительной трубки — многослойный плоский, а в среднем ее отделе — однослойный призматический. По отношению к эпителию пищеварительные железы расположены либоэндоэпителиально (например, бокаловидные клетки в кишечнике), либоэкзоэпителиально: в собственной пластинке слизистой оболочки (пищевод, желудок) и в подслизистой основе (пищевод, двенадцатиперстная кишка) или же за пределами пищеварительного канала (печень, поджелудочная железа). Собственная пластинка слизистой оболочки лежит под эпителием, отделена от него базальной мембраной и представлена рыхлой волокнистой соединительной тканью. Здесь находятся кровеносные и лимфатические сосуды, нервные элементы, скопления лимфоидной ткани. В некоторых отделах (e.g. пищевод, желудок) здесь могут располагаться простые железы. Мышечная пластинка слизистой оболочки расположена на границе с подслизистой основой и состоит из 1-3 слоев, образованных гладкими мышечными клетками. В некоторых отделах (язык, десны) гладкие мышечные клетки отсутствуют. Рельеф слизистой оболочки на протяжении всего пищеварительного канала неоднороден. Поверхность ее может быть гладкой (губы, щеки), образовывать углубления (ямочки в желудке, крипты в кишечнике), складки (во всех отделах), ворсинки (в тонкой кишке). Рельеф слизистой зависит от мышечной пластинки слизистой, а также от степени выраженности подслизистой основы. Вопрос 4. Основные зачатки и развитие лица, неба, языка. |