II. Сравнительная характеристика волокон
|
Красные
мышечные волокна
(волокна I, или
медленного типа)
|
Белые
мышечные волокна
(волокна II, или
быстрого типа)
|
Функцио-
нальные способ-
ности
|
Красные волокна способны к
не очень интенсивной,
но длительной работе.
|
Белые волокна пособны к
интенсивной,
но кратковременной работе.
|
Связь с физичес-
кими возмож-
ностями
|
Такие волокна преобладают, в частности, у стайеров - бегунов на длительные дистанции.
|
Эти волокна преобладают у спринтеров - бегунов на короткие дистанции.
|
Источ-
ник
энергии
|
Энергия получается за счёт аэробного (окислительного) распада энергетических субстратов (глюкозы, жирных кислот) до СО2 и Н2О.
|
Преобладает анаэробный (не требующий О2) распад гликогена или глюкозы до молочной кислоты.
|
а) Как видно, функциональные способности волокон связаны со способом извлечения энергии из питательных веществ - аэробным или анаэробным.
б) В свою очередь, эти общие характеристики волокон связаны с содержанием и активностью в них конкретных веществ и ферментов.
|
Мио-
глобин
|
а) В волокнах велико содержание миоглобина - белка, запасающего О2 .
б) Отсюда - красный цвет волокон (из-за наличия в миоглобине такого же пигмента, как в Hb, - гема).
|
а) Содержание миоглобина - низкое.
б) Отсюда - светлый цвет волокон.
|
Глико-
ген
|
а) В волокнах имеется гликоген,
но его запасы не очень велики.
б) Необходимости в обширных запасах нет потому, что аэробный распад веществ даёт большое количество энергии.
в) В частности, при распаде 1 молекулы глюкозы образуется 36 мол. АТФ.
|
а) Содержание гликогена - высокое.
б) Действительно, анаэробный распад 1 молекулы глюкозы даёт всего 2 мол. АТФ.
в) Поэтому для обеспечения интенсивной работы требуются большие запасы углеводов. - Которые, правда, всё равно быстро кончаются.
|
Липид-
ные включе-
ния
|
Зато в красных волокнах сравнительно многочисленны липидные включения.
|
Липидные включения невелики или вообще отсутствуют - поскольку жиры анаэробным способом не разрушаются.
|
СДГ
|
Высока активность ферментов окисления - в т.ч. сукцинатдегидрогеназы (СДГ) - фермента цикла Кребса.
Этот цикл завершает аэробный распад большинства веществ и проходит в митохондриях.
|
а) Активность СДГ, как и других митохондриальных ферментов - низкая.
б) Все ферменты анаэробного распада глюкозы содержатся в гиалоплазме (т.е. вне митохондрий).
|
АТФаза
|
Наконец, скорость распада АТФ (АТФазная активность) - относительно небольшая.
|
АТФазная активность - выше, чем в красных мышечных волокнах.
|
В мышечных волокнах распад АТФ происходит, в первую очередь, при сокращении (за счёт АТФазной активности миозиновых головок).
Поэтому скорость распада АТФ показывает, с какой скоростью может совершаться мышечная работа.
|

11.2.3.2. Препараты с гистохимической реакцией
на гликоген, АТФазу и СДГ
I. Особенности приготовления препаратов
Чтобы убедиться в том, что скелетная мышца содержит мышечные волокна разных типов, необходимо поставить соответствующие гистохимические реакции (п. 1.1.4) – в том числе на активность ряда ферментов.
В этом случае приготовление препаратов имеет существенные особенности.
а) Взятый образец ткани
сразу не фиксируют (тогда все ферменты просто потеряли бы свою активность),
а подвергают глубокому замораживанию.
б) После замораживания приготовляют срезы.
в) Срезы размораживают и помещают в инкубационную среду – с таким составом,
чтобы прошла изучаемая реакция
и чтобы один из её продуктов дал окрашенное соединение.
г) Затем срезы промывают, фиксируют и заключают в консервирующую среду.
|
II. Просмотр препаратов
Гликоген
|
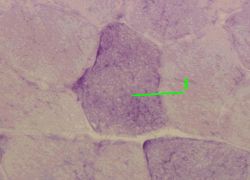
3. Препарат - гликоген в скелетных мышечных волокнах. ШИК-реакция.
|
а) На снимке видны поперечно срезанные мышечные волокна.
б) В их саркоплазме находятся глыбки гликогена (1), окрашенные в тёмно-малиновый цвет.
в) Более светлые на снимке волокна содержат меньше гликогена:
это волокна I типа ("красные").
|
Полный размер
|
АТФаза
|
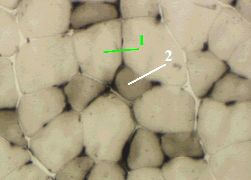
4. Препарат - типы мышечных волокон. Реакция на АТФазную активность.
|
Здесь тоже наблюдается подразделение мышечных волокон на 2 типа.
Волокна I типа (1) и при данной окраске являются более светлыми,
что означает меньшую АТФазную активность.
Волокна II типа (2) - более тёмные:
АТФаза быстрого типа.
|
Полный размер
|
СДГ
|
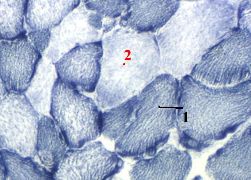
5. Препарат - типы мышечных волокон. Реакция на сукцинатдегидрогеназу.
|
1. а) Фермент СДГ содержится в митохондриях.
б) Поэтому гранулы проявления их активности, окрашенные в тёмно-синий цвет, маркируют расположение митохондрий.
2. Вновь видны различия между волокнами. -
|
Полный размер
|
Волокна I типа (1) теперь являются более тёмными - из-за большей активности СДГ,
а волокна II типа (2) - более светлыми .
|
11.3. Сердечная поперечнополосатая мышечная ткань
Как уже отмечалось, сердечная мышечная ткань развивается изутолщения висцерального листка спланхнотома – миоэпикардиальной пластинки.
|
11.3.1. Клеточная организация ткани
11.3.1.1. Типичные кардиомиоциты и функциональные волокна
9. Препарат - срез миокарда. Окраска железным гематоксилином.
|
Объеди-
нение в функцио-
нальные волокна
|
а) Основной элемент сердечной мышечной ткани – типичные кардиомиоциты (слово “типичные”часто опускают).
б) Этоклетки цилиндрической формы, которые
стыкуются друг с другом своими основаниями,
образуя функциональные волокна.
|

Полный размер
|
Связи между "волок-
нами"
|
"Волокна" связаны многочисленными анастомозами – за счёт того, что в этих участках кардиомиоциты
на концах раздвоены и
контактируют с клетками сразу двух "волокон".
|
Базаль-
ная мембрана
|
а) Функциональные волокна окружены базальной мембраной.
б) Таким образом, последняя
покрывает лишь боковые поверхности кардиомиоцитов,
но не заходит на их основания (торцевые поверхности).
|
Размеры
|
а) Диаметр клеток (а значит, и диаметр "волокон") – около 20 мкм. Это
существенно меньше диаметра истинных волокон скелетной мышечной ткани (примерно 50-70 мкм; см. выше).
б) Длина кардиомиоцитов – примерно 100 мкм.
|
11.3.1.2. Вставочные диски
Опреде-
ление
|
а) Места контактов соседних кардиомиоцитов в функциональных волокнах называются вставочными дисками (2).
б) На световых препаратах они выглядят как тонкие и чёткие поперечные тёмные полоски в "волокнах".
|
|
Составле-
ние с поперечной исчерчен-
ностью
|
а) Не надо путать эти полосы с более мелкой поперечной исчерченностью (1) кардиомиоцитов, обусловленной исчерченностью миофибрилл (т.е. чередованием I- и А-дисков).
б) Уже отмечалось, что период саркомерной организации миофибрилл составляет в покое 2,3мкм.
в) Расстояние же между соседними вставочными дисками - это длина кардиомиоцита, т.е. 100 мкм (см. выше).
г) Отсюда следует, что между соседними чёткими чёрточками (вставочными дисками) находится несколько десятков (40-50) еле заметных чёрточек - А-дисков.
|
Межкле-
точные контакты
|
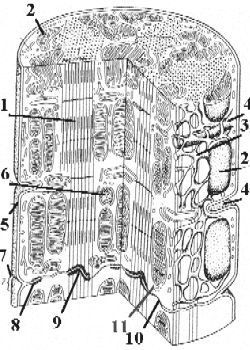
В области вставочных дисков (9) между кардиомиоцитами существуют контакты трёх видов (раздел 2.3):
интердигитации – пальцевидные впячивания клеток друг в друга;
десмосомы (10) – контакты, обеспечивающие более прочное сцеплениеклеток;
нексусы (11) – контакты, пронизанные гидрофильными каналами и потому обеспечивающие электрическую и метаболическую связь между кардиомиоцитами.
|
Схема - строение кардиомиоцитов и вставочных дисков.
|
11.3.1.3. Дополнительные клеточные элементы
Отсутствие камбия
|
а) Миосателлитов или иных камбиальных клеток в сердечной мышечной ткани нет.
б) А сами кардиомиоциты утрачивают способность делиться к моменту рождения ребёнка или в первые месяцы жизни.
в) Поэтому при регенерации
новые кардиомиоциты и функциональные волокна не образуются,
а происходит только гипертрофия (увеличение объёма) сохранившихся клеток.
|
Разновид-
ности кардио-
миоцитов
|
а) Однако кроме типичных (сократительных, или рабочих) кардиомиоцитов, в сердце присутствуют и другие их разновидности:
секреторные, а также
атипичные кардиомиоциты(составляющие проводящую систему сердца).
б) Эти клетки значительно отличаются по своей морфологии, метаболизму, функции и будут рассмотрены позже – в теме 19.
|
11.3.2. Строение типичных кардиомиоцитов
11.3.2.1. Органеллы кардиомиоцитов
Мио-
фибриллы
|
а) Миофибриллы (1) имеют такую же организацию, как и в скелетной мышечной ткани.
б) Но их относительное содержание (по сравнению с той же тканью) меньше: они занимают лишь 40% объёма кардиомиоцитов.
в) В области вставочных дисков миофибриллы прикрепляются к плазмолемме (2).
|
|
Ядра
|
а) В клетке присутствует 1-2 ядра – как правило, полиплоидные.
б) Из-за относительно небольшого содержания миофибрилл, ядра не оттесняются к периферии, а
остаются в центре клетки.
|
Мембран-
ные системы
|
В типичных кардиомиоцитах, как и в мышечных волокнах скелетных мышц, имеются специальные мембранные системы:
Т-трубочки (4) – глубокие впячивания плазмолеммы, идущие вокруг миофибрилл,
и L-система (L-канальцы и терминальные цистерны) (3) – производное саркоплазматического ретикулума.
|
|
Прочее
|
Также в кардиомиоцитах велико содержание митохондрий(5).
|
11.3.2.2. Гистохимические особенности кардиомиоцитов
Резервные структуры
|
а) Наряду с митохондриями, в кардиомиоцитах
много миоглобинаи липидных капель.
б) Гранул же гликогена относительно мало.
|
Тип обмена
|
а) Такое соотношение резервов отражает тот факт, что в кардиомиоцитах реализуется
аэробный способ разрушения питательных веществ.
б) В этом отношении кардиомиоциты
подобны краснымволокнам скелетных мышц.
|
Продукты "питания" кардио-
миоцитов
|
Субстратами окисления служат следующие вещества:
в первую очередь, жирные кислоты,
кроме того, – продукты распада последних – т.н. кетоновые тела (ацетоацетат и др.), поступающие из печени;
а в период интенсивной мышечной нагрузки – ещё и лактат, поступающий из скелетных мышц, и глюкоза, потребление которой возрастает.
|
Липо-
фусцин
|
Как отмечалось в п.3.2.4, с возрастом в кардиомиоцитах накапливается пигмент старения липофусцин – продукт неполного переваривания веществ в телолизосомах.
|
11.3.2.3. Резюме: различия скелетной и сердечной
мышечных тканей
а) Таким образом, сердечная мышечная ткань имеет
и выраженные черты сходства со скелетной мышечной тканью,
и не менее характерные отличия.
б) Последние кратко суммированы в следующей таблице.
| |
 Скачать 1.48 Mb.
Скачать 1.48 Mb.