Основы строения головного и спинного мозга. Учебнометодическое пособие для студентов педиатрического и лечебного факультетов
 Скачать 2.31 Mb. Скачать 2.31 Mb.
|

|
). Этими отверстиями четвертый желудочек сообщается с подпаутинным пространством. Рельеф ромбовидной ямки (см. рис.9). Из боковых углублений, recessus lateralis, к середине ромбовидной ямки тянутся белые мозговые (слуховые) полоски, striae medullaris (acusticae), разделяющие ее на верхний и нижний треугольники. Продольная срединная борозда, sulcus medianus, разделяет ромбовидную ямку на две симметричные половины. По обеим сторонам от срединной борозды располагается срединное возвышение, eminentia medialis, наружной границей которого является пограничная борозда, sulcus limitans. Сверху и снизу пограничная борозда заканчивается углублениями - верхней и нижней ямками, fovea superior et inferior. Верхняя ямка соответствует месту расположения двигательного ядра тройничного нерва, а снаружи от него проецируется чувствительное ядро этой пары. Здесь находится окрашенное в синеватый цвет голубое пятно, locus caeruleus, которое соответствует латеральной границе ромбовидной ямки. Специфический цвет пятна обусловлен нервными клетками, в цитоплазме которых имеются включения нейромеланина. В верхней части ромбовидной ямки срединное возвышение расширяется и сильнее вдается в полость желудочка в виде округлой формы лицевого бугорка, colliculus facialis. Он образован подлежащим двигательным ядром отводящего нерва и огибающими его волокнами лицевого нерва. Книзу срединное возвышение суживается, принимает треугольную форму и получает название треугольника подъязычного нерва, trigonum nervi hypoglossi. Он определяет локализацию ядра XII пары. Снаружи от него лежит сероватого цвета треугольное поле, известное под названием серого крыла, ala cinerea, или треугольника блуждающего нерва, trigonum nervi vagi. Сюда проецируется дорсальное ядро X пары. Кнаружи и выше дорсального ядра лежит проекционное поле ядра одиночного пути (общее ядро VII, IX и X нервов). Рядом с ним, параллельно срединной борозде проецируется спинномозговое ядро тройничного нерва. Наружный отдел ромбовидной ямки, расположенный между пограничной бороздой и входом в боковой карман, называют вестибулярным полем, area vestibularis. Под ним лежат ядра предцверно-улиткового нерва. Средний мозг Состав среднего мозга. Mesencephalon представляет собой наименьшую из частей головного мозга. Его дорсальная часть представлена крышей среднего мозга, tectum mesencephalic которая также называется пластинкой четверохолмия, lamina quadrigemina. Вентральная часть образована ножками мозга, pedunculi cerebri. Полость среднего мозга, aquaeductus cerebri, соединяет III и IV желудочки. Крыша среднего мозга Внешнее строение. Пластинка четверохолмия, tectum mesencephalic простирается от основания эпифиза до переднего конца верхнего мозгового паруса (см. рис.9). Она образована верхними бугорками, colliculi superiores, и нижними бугорками, colliculi inferiores. Между верхними бугорками в trigonum subpineale располагается шишковидная железа. От крыши среднего мозга к верхнему мозговому парусу тянутся две уздечки, frenula veli medullaris anterioris. Сбоку от уздечки с каждой стороны выходит nervus trochlears. Бугорки снаружи переходят в ручки холмиков. Соответственно от верхних бугорков отходит верхняя ручка, brachium colliculi superior is, которая направляется к латеральному коленчатому телу. От нижних бугорков отходит нижняя ручка, brachium colliculi inferior is, к медиальному коленчатому телу. Внутреннее строение. В верхних бугорках четверохолмия находится один из подкорковых зрительных центров. Бугорки состоят из чередующихся доев серого и белого вещества. Эти слои обычно подразделяют на поверхностные и глубокие. Поверхностные слои реагируют на зрительный стимул, получая сенсорную информацию из сетчатки и передавая ее в кору через латеральное коленчатое тело и подушку зрительного бугра. От глубоких слоев начинаются нисходящие проводящие пути, связанные с движением головы и глаз: tr. tectospinalis, tr, tectobulbaris, tr. tectopontinus. Верхние бугорки выполняют функцию рефлекторных центров, которые определяют положение головы и глаз в ответ на зрительные и слуховые раздражения. Они обеспечивают слежение за направлением движения объекта в зрительном поле, зрительную ориентацию, наблюдение. Нижние бугорки четверохолмия - основной подкорковый центр слуха, который состоит из трех ядер: центрального, nucL centralis, околоцентрального, nucl pericentralis и наружного, nucl. extemus. Центральное и околоцентральное ядра нижних холмиков передают информацию в медиальное коленчатое тело, откуда она поступает в слуховой центр коры больших полушарий. Наружное ядро обеспечивает акустическо-двигательную реакцию за счет связей с глубокими слоями верхних бугорков четверохолмия, откуда начинаются нисходящие проводящие пути. Ножки мозга Внешнее строение. Ножки мозга, pedunculi cerebri, идут от верхнего края моста кверху и в латеральном направлении, а затем погружаются в толщу полушарий большого мозга. В области входа в полушария через ножки перекидываются зрительные тракты. Между ножками мозга находится fossa interpeduncular (Tarini% дно которой образовано задним продырявленным веществом. На медиальной поверхности ножек мозга лежит глазодвигательная борозда, sulcus oculomotorius место выхода волокон одноименного нерва. Внутреннее строение (рис.13). На поперечных разрезах среднего мозга обнаруживается серовато-черное вещество, substantia nigra (Sommering), которое разделяет ножки на основание, basis pedunculi cere bra! is, и покрышку, tegmentum. Снаружи граница основания и покрышки проходит по sulcus nervi oculomotorii. Водопровод мозга на поперечном разрезе имеет вид треугольника с основанием, направленным вверх, и верхушкой - вниз. В основании ножек мозга проходят нисходящие проводящие пути: Tr. corticospinalis часть пирамидного пути к спинному мозгу. Tr. corticonuclearis - часть пирамидного пути к двигательным ядрам черепных нервов. Tr. corticopontinus, корково-мостовой путь, который включает fibrae frontopontinae, которые начинаются в лобной доле и занимают медиальную 1/6 часть основания ножки мозга и fibrae parietotemporopontinae - из теменной и височной долей, расположенные в латеральной части ножек. Серое вещество покрышки ножек мозга: Центральное серое вещество, substantia grisea centralis - окружает водопровод мозга. Содержит несколько ядерных групп, большинство из которых входит в состав лимбической системы мозга. Ядра черепных нервов III, IV и среднемозговое ядро V пары черепных нервов. Глазодвигательный нерв (III) имеет двигательное соматическое ядро, расположенное кпереди от центрального серого вещества и вегетативное парасимпатическое ядро (Якубовича, Edinger-Westphal). Двигательное ядро блокового нерва находится в центральном сером веществе, ниже ядра III пары. Ядра III пары располагаются на уровне верхних, а ядра IV пары - нижних бугорков четверохолмия. 3. Добавочные ядра глазодвигательного комплекса: Nucleus interstitialis (Cajal), промежуточное ядро Кахаля - лежит снаружи от ядра глазодвигательного нерва, от которого отделяется медиальным продольным пучком. Предполагается его участие в медленных вращательных и вертикальных движениях глазных яблок, в контроле движений и позы головы. • Nucleus centralis (Perlia) -расположенная срединно группа клеток. Предполагается ее участие в реакции конвергенции. Красное ядро, nucleus ruber, расположено между substantia nigra и центральным серым веществом. В его состав входят нейроны с железосодержащим пигментом, на которых заканчиваются волокна от коры большого мозга, таламуса и мозжечка (fibrae dentatorubrales). От нейронов красного ядра начинается красноядерно-спинномозговой (Монакова) путь, tr. rubrospinal (Monakow), который направляется к клеткам передних рогов спинного мозга. Относится к экстралирамидной системе. Р 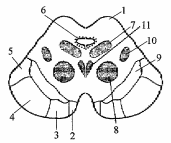 ис.13. Поперечный разрез среднего мозга: ис.13. Поперечный разрез среднего мозга:1 - colliculus superior; 2 - fibrae frontopontinae; 3 - tractus corticonuclearis; 4 - tractus corticospinalis; 5 - fibrae parietotemporopontinae; 6 - substantia grisea centralis; 7 - nuclei nervi oculomotorii; 8 - nucleus ruber; 9 - substantia nigra; 10 - lemniscus medialis; 11 - fasciculus longitudinalis medialis Черное вещество, substantia nigra, входит в состав стриопаллидарной системы, получая наибольшее число волокон от хвостатого ядра, fibrae strionigrales. Ретикулярная формация, formatio reticularis, лежит дорсальнее и латеральнее красного ядра. Белое вещество покрышки ножек мозга: Латеральная петля, lemniscus lateralis проводящий путь слухового анализатора. Медиальная петля, lemniscus medialis, образована аксонами вторых нейронов проводящих путей проприоцептивной чувствительности коркового направления (Голля и Бурдаха). Спинномозговая петля, lemniscus spinalis, образована аксонами вторых нейронов спинно-таламического пути. Проходит в непосредственной близости от медиальной петли. Покрышечно-спинномозговой и покрышечно-бульбарный пути, tr. tectospinal et tr. tectobulbaris, начинаются от нейронов глубоких слоев верхних бугорков четверохолмия. Направляются вентро-медиально и, огибая центральное серое вещество, переходят на противоположную сторону (decussatio tegmentalis dorsalis). Tr. tectospinalis заканчивается на мотонейронах передних рогов серого вещества шейных сегментов спинного мозга. Tr. tectobulbaris направляется к ядрам глазодвигательных нервов. Обеспечивают рефлекторную двигательную реакцию на зрительные и слуховые раздражения. Входят в состав экстрапирамидной системы. Красноядерно-спинномозговой путь, tr. rubrospinal, начинается от нейронов красного ядра. Волокна пути переходят на противоположную сторону (decussatio tegmentalis ventralis) и спускаются в составе бокового канатика к мотонейронам передних рогов серого вещества спинного мозга. Входит в состав экстрапирамидной системы. Медиальный продольный пучок, fasciculus longitudinalis medialis, соединяет ядра глазодвигательных нервов и вестибулярные ядра с мотонейронами передних рогов, иннервирующими мышцы шеи. Обеспечивает сочетанные движения головы и глаз. Ретикулярная формация мозгового ствола Ретикулярная формация, forrnatio (substantia) reticularis, образована переплетением волокон, идущих в различных направлениях, и расположенными между ними нервными клетками, которые образуют отдельные клеточные группы (ядра ретикулярной формации). Ретикулярная формация лежит в дорсальной части продолговатого мозга, моста, ножек мозга и простирается в каудальную часть промежуточного мозга. Ее рассматривают как ростральное продолжение межнейронных сетей спинного мозга, которые становятся значительно более интенсивными. Аксоны нервных клеток ретикулярной формации опускаются в спинной мозг, восходят к таламическому и гипоталамическому отделам промежуточного мозга, к коре полушарий. Выделяют 4-е области ретикулярной формации мозгового ствола: Срединная область, zona mediana. Околосрединная область, zona paramediana - аксоны нейронов этой локализации имеют преимущественно мозжечковое направление. Медиальная область, zona medialis, которую так же называют гигантоклеточной, является источником восходящих и нисходящих волокон различных направлений (эфферентное поле ретикулярной формации). Латеральная область, zona lateralis (мелкоклеточная), представляет собой афферентное (сенсорное) поле, так как здесь заканчиваются коллатерали всех чувствительных путей. Аксоны нейронов этой локализации направляются в медиальную область ретикулярной формации. Ретикулярная формация представляет собой полифункциональную структуру. Она является интегративньш центром ствола во многом определяющим направление и интенсивность потоков сенсорной информации к высшим отделам мозга, а также степень нисходящих влияний на активность спинальных и стволовых нейронов. Ретикулярная формация участвует в регуляции рефлекторной двигательной активности. Из эфферентного поля ретикулярной формации начинаются два ретикулоспинальных пути: tractus reticulospinalis medialis, который содержит волокна из области моста, и tractus reticulospinalis lateralis, начинающийся из ядерной зоны продолговатого мозга. Волокна медиального пути не переходят на противоположную сторону. В состав латерального тракта входят как перекрещенные, так и неперекрещенные волокна. Оба пути заканчиваются на интернейронах серого вещества спинного мозга. Установлено, что волокна от продолговатого мозга возбуждают альфа- и гамма-мотонейроны сгибателей и вызывают торможение мотонейронов разгибателей., тогда как волокна из области моста оказывают обратное действие. Таким образом, ретикулярная формация может как облегчать, так и подавлять рефлекторную двигательную активность. Ядра ретикулярной формации образуют основные «центры жизнеобеспечения» ствола, такие как дыхательный и сосудодвигательный. Дыхательный центр продолговатого мозга окружает nucl. solitarius et nucl. ambiguus. Он состоит из центра вдоха, расположенного в гигантоклеточной области ретикулярной формации, и центра выдоха, занимающего мелкоклеточную область. В ретикулярной формации моста располагаются «пневмотаксический» центр (центр регуляции внешнего дыхания) и центр «апноэ» (задержки дыхания). «Прессорный» и «депрессорный» сосудодвигательные центры обеспечивают рефлекторную регуляцию кровяного давления, «акселераторный» и «ингибиторный» центры - частоты и силы сердечных сокращений. Ретикулярная формация участвует в модуляции болевой чувствительности. Установлено, что стимуляция определенных областей сетчатой субстанции может заметно уменьшить или прервать афферентную импульсацию Ретикулярная формация обладает выраженным активизирующим действием на активность нейронов коры больших полушарий. Восходящие пути из ретикулярной формации охватывают широкие корковые поля. Они получили название «второй восходящей системы» или «восходящей ретикулярной активирующей системы», в отличие от «первой восходящей системы», включающей так называемые «лемнисковые» пути. Восходящая ретикулярная активирующая система представляет собой полисинаптические пути проведения в кору импульсов неспецифической природы, связанных с «пробуждением и побуждением». Эти пути начинаются в эфферентной области ретикулярной формации. Среди них главный восходящий путь ствола - tractus tegmentalis centralis. Этот путь проходит через ростральные внутрипластинчатые ядра таламуса. Непрерывный поток афферентной импульсации из ретикулярной формации создает оптимальной состояние коры для осознанного восприятия специфических чувствительных сигналов, передаваемых классическими («лемнисковыми») проводящими путями. Промежуточный мозг В состав промежуточного мозга входят область зрительного бугра, thalamencephalon, и подталамическая область, hypothalamus, разделеные sulcus hypothalamics, которая соединяет foramen interventriculare и aqueductus cerebri. Полостью промежуточного мозга является непарный третий желудочек, ventriculus tertius, лежащий в срединной плоскости. Thalamencephalon, в свою очередь, слагается из зрительного бугра, thalamus; надталамической области, epithalamus, и заталамической области, metathalamus. Зрительный бугор Внешнее строение. Зрительный бугор, thalamus, имеет свободные дорсальную и медиальную поверхности, которые разделены мозговой полоской, stria medullaris (см. рис.9). На границе зрительного бугра и хвостатого ядра находится терминальная, полоска, stria terminalis. На дорсальной поверхности определяются передний бугорок, tuberculum anterius thalami, и подушка, pulvinar (задняя утолщенная часть таламуса). Дорсальная поверхность покрыта тонким слоем белого вещества, который называется поясной слой, stratum zonale. Медиальные поверхности соединяются между собой с помощью межталамического сращения, adhesio interthalamica. Вентральная поверхность зрительного бугра прилежит к hypothalamus, а латеральная граничит с внутренней капсулой. Внутреннее строение. С позиций функциональной анатомии таламус обычно называют «коллектором чувствительности» или «воротами в кору», так как все афферентные пути коркового направления проходят через зрительный бугор и имеют здесь свои, так называемые «ядра переключения». Связи этих ядер с корой в большинстве случаев являются реципрокными (двухсторонними). По функции все ядра таламуса подразделяются на 4 группы: Специфические ядра переключения для афферентных систем. В этих ядрах происходит передача определенной сенсорной информации (общая чувствительность, зрительная, слуховая). Их афферентные проекции соответствуют сенсорным областям коры. Ядра неспецифической системы. Эти ядра связаны с ретикулярной формацией ствола мозга и получают информацию от всех органов чувств. Ядра с преимущественно моторными функциями являются ассоциативным звеном, которое обеспечивает связь мозжечка и базальных ядер конечного мозга с двигательными зонами коры больших полушарий. Ядра с ассоциативными функциями. Они связаны с ассоциативными областями коры, которые принимают участие в высших интегративных процессах головного мозга. По топографическому принципу ядра таламуса объединены в три основные ядерные группы: Передние ядра, nuclei anteriores; Медиальные ядра, nuclei mediales; Латеральные ядра, nuclei laterales thalami. Эти группы отделены друг от друга внутренней мозговой пластинкой, laminae medullares interna. Передняя группа состоит из переднемедиального, nucl anteromedialis, переднедорсального, nucl. anterodor salts, и передневентрального, nucl anteroventralis, ядер. Эти ядра связаны с лимбической корой и с подкорковыми лимбическими центрами ствола мозга, поэтому их называют «лимбическимй ядрами» зрительного бугра. В эксперименте на лабораторных животных показано, что раздражение или разрушение данной области вызывает состояние «настороженности и нападения». Эти ядра также вовлечены в процессы обучения и памяти. Медиальная группа представлена дорсальным медиальным ядром, nucl medialis dorsalis, которое связано со всеми корковыми полями лобной доли кпереди от предцентральной извилины, включая её латеральную, медиальную и базальную поверхности. Так же как и передняя группа ядер принадлежит к лимбической системе, но при его экспериментальном разрушении развивается эмоциональная обедненность ответной реакции. Медиальная группа ядер также связана с познавательными функциями памяти и обучения. Латеральная группа ядер делится на дорсальную и вентральную. К дорсальной группе относятся дорсальное латеральное ядро, nucl. lateralis dorsalis, заднее латеральное ядро, nucl lateralis posterior, и ядра подушки. Все они относятся к ассоциативным ядрам зрительного бугра. В частности, корковая проекция подушки - ассоциативная зрительная кора. К вентральным ядрам относятся: переднее вентральное ядро, nucl ventralis anterior, (ассоциативное), латеральное вентральное ядро, nucl. ventralis lateralis, (преимущественно двигательные функции), задние вентральные ядра, nuclei centrales posteriores. К задним вентральным ядрам относятся заднелатеральное, nucl ventralis posterolateralis (VPL), и заднемедиальное, nucl ventralis posteromedialis (VPM) ядра. В VPL заканчиваются медиальная и спинальная петли, которые являются проводниками проприоцептивной и экстероцептивной чувствительности от туловища и конечностей. В VPM заканчивается тройничная петля - проводник всех видов чувствительности от области лица. Для этих ядер характерна соматотопическая организация, т.е. каждое рецепторное поле проецируется в определенную область ядра. Внутри мозговой пластинки также имеются ядра, среди которых наиболее выражено медиальное центральное. Они относятся к ядрам неспецифической системы. Внутрипластинчатые ядра зрительного бугра имеют связи с ретикулярной формацией и черным веществом ножек мозга. Эпиталамус Epithalamus примыкает- к заднему концу зрительных бугров (см. рис.9). Stria medullaris каудально расширяется, образуя треугольное поле, trigonum habenulae, от которого по направлению к средней линии тянется тонкий волокнистый тяж - поводок, habenula, образующий вместе с таким же поводком противоположной стороны спайку, commissura habenularum. Спайка поводков соединяется с шишковидной железой, epiphysis (corpus pineale). Все вышеперечисленные образования и составляют надталамическую область. В основании эпифиза можно обнаружить слепо замкнутое выпячивание третьего желудочка, recessus pinealis. Под ним находится, вдающийся в желудочек, пучок поперечно идущих волокон - задняя мозговая спайка, commissura cerebri posterior. Метаталамус Заталамическую область, metathalamus, образуют латеральное и медиальное коленчатые тела, corpus geniculatum laterale et mediale, лежащие позади зрительного бугра (см. рис.8). Латеральное коленчатое тело является основным подкорковым центром зрительного, а медиальное - слухового анализаторов. Гипоталамус В состав гипоталамической области входят образования различного генеза. Они занимают вентральную часть промежуточного мозга и располагаются ниже подбугорной борозды, sulcus hypothalamics. В ростро- каудальном направлении выделяют: Pars optica hypothalami, которая простирается от lamina terminalis до задней поверхности зрительного перекреста. Pars tuberalis, в состав которой входят tuber cinereum, infundibulum et hypophysis. Pars mammilaris - область сосцевидных тел. При этом каждая область включает несколько ядерных групп (рис. 14). В pars optica описаны: Преоптическая область, area preoptica - обеспечивает регуляцию секреции гонадотропных гормонов передней доли гипофиза, выделяя гормон-регулирующие факторы, Паравентрикулярное ядро, nucl paraventricular, и супраоптическое ядро, nucl supraopticus образованы телами крупноклеточных эндокринных нейронов. В этих ядрах образуются висцеротропные гормоны - окситоцин и вазопрессин. Супрахиазматическое ядро, nucl suprachiasmaticus - принимает участие в управлении циркадными ритмами. Имеет прямую и непрямую связь с сетчаткой. В pars tuberalis расположены: 1. Дугообразное ядро, nucl arcuatus - образовано телами мелкоклеточных эндокринных нейронов. В ядре происходит образование гормон-регулируюгцих факторов, определяющих секреторную активность клеток аденогипофиза. 2. Дорсомедиальное ядро, nucl. dorsomedialis, и вентромедиальное ядро, nucl. ventromedial - являются центром регуляции парасимпатическим отделом вегетативной нервной системы. Эти ядра имеют связь со стволовыми и сакральным парасимпатическими центрами. В pars mamillaris находятся: Заднее ядро, nucl. posterior - является центром регуляции симпатическим отделом вегетативной нервной системы. Оно связано с боковым промежуточным ядром спинного мозга. Ядра сосцевидного тела, nuclei corporis mamillaris medialis et lateralis - являются частью обонятельного мозга. Таким образом, гипоталамус является главным подкорковым центром регуляции вегетативно-висцеральных и эндокринных функций. Р 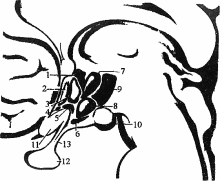 ис.14. Схема строения гипоталамической области промежуточного мозга: ис.14. Схема строения гипоталамической области промежуточного мозга:1-nucl. paraventricularis; 2-area preoptica; 3-nucl. anterior; 4»nucl. suprachiasmaticus; 5-nucl. supraopticus; 6-nucl. arcuatus; 7- nucl dorsomedialis; 8-rtucl. ventromedialis; 9-nuci posterior; 10-nuclei corporis mammiiaris medialis et lateralis; 11-chiasma opticum; 12- hypophysis; 13-infundibulum Третий желудочек Полость промежуточного мозга, ventriculus tertius, через foramen interventriculare сообщается с боковыми желудочками, а через aquaeductus cerebri с четвертым. Стенки третьего желудочка: Передняя: lamina terminalis, commissura cerebri anterior, columnae fornicis. Задняя: commissura habenularum, corpus pineale, commissura cerebri posterior. Боковые: facei mediates thalami. Дно: regio hypothalamica. Крыша: lamina epithelial is chorioidea и вторично tela chorioidea ventriculi tertii, fornix et corpus callosum. Сосудистая покрышка третьего желудочка, tela chorioidea ventriculi tertii, состоит из двух листков, производных мягкой мозговой оболочки, между которыми просматриваются внутренние мозговые вены, vv. cerebri internae. В результате их слияния образуется большая мозговая вена, v. cerebri magna (Galeni). Нижний листок tela chorioidea образует ворсинки, которые содержат сосуды и тянутся двумя рядами параллельно срединной плоскости, провисая в полость третьего желудочка и образуя его сосудистое сплетение, plexus chorioideus ventriculi tertii. Co стороны полости желудочка сосудистое сплетение покрыто эпителиальной пластинкой, lamina epithelialis chorioidea (крыша третьего желудочка). Конечный мозг Конечный мозг (telencephalon) представлен двумя полушариями, hemispheria cerebri. В состав конечного мозга входят плащ, pallium - кора большого мозга; базальные ядра, nuclei basales; обонятельный мозг, rhinencephalon и белое вещество полушарий. Полостью конечного мозга являются боковые желудочки, ventriculi laterales. Кора большого мозга В каждом полушарии различают три поверхности: верхнелатеральную, fades superolateralis, нижнюю (базальную), fades inferior, и медиальную, fades medialis; три полюса: лобный, polus frontalis, затылочный, polus occipitalis, и височный, polus temporalis, и три края: верхний, margo superior, нижний, margo inferior, и наружный, margo lateralis. Постоянными, глубокими щелями или первичными бороздами полушария разделены на 5 долей: лобную, теменную, височную, затылочную и островок. Вторичные и третичные борозды определяют рельеф этих долей. Между бороздами находятся извилины. Такое складчатое строение увеличивает площадь коры до 2500 см2, при этом около 30% серого вещества находится на поверхности извилин и почти 70% в глубине борозд. Толщина коры в различных областях колеблется от 1,3 мм до 4,5 мм. В её состав входит 109 - Ю10 нейронов и еще большее число глиальных клеток, а также нервные волокна (афферентные, эфферентные, ассоциативные и комиссурапъные). В пределах коры наблюдается чередование слоев, содержащих преимущественно тела нервных клеток, со слоями, образованными в основном их аксонами. На основании цитоархитектоники, т.е. строения, характера и плотности расположения нервных клеток, в коре с типичным строением выделяется шесть слоев (пластинок) (обозначаются римскими цифрами): 1.1- lamina molecularis (plexiformis), молекулярная пластинка, состоит из небольшого количества горизонтально ориентированных клеток (горизонтальные клетки Кахаля), отростки которых не выходят за пределы коры, а также густой сети, образованной дендритами и аксонами клеток более глубоко расположенных слоев. II - lamina granulans externa, наружная зернистая пластинка, в ее состав входят тела звездчатых нейронов и мелких пирамидных клеток, а также сеть тонких нервных волокон. Ill - lamina pyramidalis externa, наружная пирамидная пластинка, содержит тела пирамидных нейронов среднего размера, отростки которых не образуют длинных проводящих путей. IV - lamina granulans interna, внутренняя зернистая пластинка, состоит из плотно расположенных мелких пирамидных и звездчатых нейронов. В этом слое заканчивается основная часть таламических нервных волокон. V - lamina pyramidalis interna (ganglionaris), внутренняя пирамидная пластинка, образована крупными пирамидными клетками, а в области двигательной зоны коры - гигантскими пирамидными клетками. Из пятого слоя начинаются корково-ядерные и корково-спинномозговые пути. VI - lamina multiformis, мультиформная пластинка, состоит из разнообразных по форме нейронов. Такое шестислойное строение имеет 90% коры головного мозга. В процессе филогенеза подобная кора появляется у млекопитающих, и поэтому носит название иеокортекса (новой коры). Ее также называют изокортекс вследствие однотипной организации. 10% коры занимает аллокортекс с нетипичной структурной организацией - это старая кора, archicortex, древняя кора, paleocortex, и средняя кора, mesocortex. Старая, с филогенетической точки зрения, часть коры большого мозга представлена гиппокампом и зубчатой извилиной. Она имеет только три слоя нейронов. Древняя кора соответствует области мозга, которая развивается из грушевидной доли. К средней коре относится не полностью дифференцированная кора островка. Борозды и извилины верхнелатеральной поверхности полушарий. На верхнелатеральной поверхности полушария глубокая латеральная борозда, sulcus lateralis (Sylvii), отделяет височную долю от лобной и теменной. По направлению от височного полюса к затылочному, она разделяется на три ветви: ramus anterior, идущую горизонтально вперед, ramus ascendens, направленную вертикально вверх, и более длинную ramus posterior, которая продолжается назад. От середины верхнего края полушарий косо вниз и вперед направляется центральная борозда, sulcus centralis (Rolandi), которая разделяет лобную и теменную доли (рис.16,17). Lobus frontalis Борозды лобной доли: Преддентральная борозда, sulcus precentralis, расположена кпереди от центральной борозды и параллельно ей. Верхняя лобная борозда, sulcus frontalis superior, направляется вперед от верхней части предцентральной борозды. Нижняя лобная борозда, sulcus frontalis inferior, начинается от нижней части предцентральной борозды. Средняя лобная борозда, sulcus frontalis medius - непостоянная, проходит между верхней и нижней лобными бороздами. Извилины лобной доли: Предцентральная извилина, gyrus precentralis (лежит между sulcus centralis и sulcus precentralis). Верхняя лобная извилина, gyrus frontalis superior (помещается над sulcus frontalis superior). С 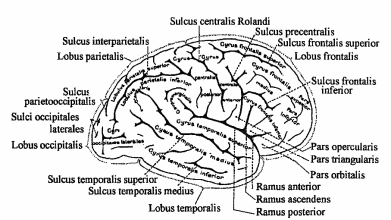 редняя лобная извилина, gyrus frontalis medius (находится между sulcus frontalis superior и sulcus frontalis inferior, и посредством sulcus frontalis medius разделяется на две части pars superior и inferior ). редняя лобная извилина, gyrus frontalis medius (находится между sulcus frontalis superior и sulcus frontalis inferior, и посредством sulcus frontalis medius разделяется на две части pars superior и inferior ).Нижняя лобная извилина, gyrus frontalis inferior (расположена под sulcus frontalis inferior). Рис. 16. Рельеф верхнелатеральной поверхности полушарий. Последняя извилина называется также извилиной Брока и разделена на три отдела: Покрышечная часть, pars operculars - между нижним концом sulcus precentralis и ramus ascendens боковой борозды. Треугольная часть, pars triangularis - между ramus ascendens и ramus anterior боковой борозды. Глазничная часть, pars orbitalis - кпереди от ramus anterior. Lobus parietalis Борозды теменной доли: Постцентральная борозда, sulcus postcentrals, проходит позади sulcus centralis и параллельно ей. Внутритеменкая борозда, sulcus intraparietalis, начинается от постцентральной борозды и направляется дугообразно назад. Извилины теменной доли: Постцентральная извилина, gyrus postcentralis, лежит позади sulcus centralis. Верхняя теменная долька, lobulus parietalis superior, располагается над sulcus intraparietalis. Нижняя теменная долька, lobulus parietalis inferior, лежит под sulcus intraparietalis. Нижняя теменная долька состоит из двух отдельных извилин: надкраевой, gyrus supramarginalis, и угловой, gyrus angularis. Gyrus supramarginalis охватывает восходящую конечную ветвь боковой борозды. Gyrus angularis располагается над проксимальным концом верхней височной борозды. Lobus temporalis |