био расписанные билеты мои. Уровни организации жизни. Проявление главных свойств жизни на разных уровнях ее организации
 Скачать 0.93 Mb. Скачать 0.93 Mb.
|

|
Цели создания ГМО Продовольственная и сельскохозяйственная организация ООН рассматривает использование методов генетической инженерии для создания трансгенных сортов растений либо других организмов как неотъемлемую часть сельскохозяйственной биотехнологии. Прямой перенос генов, отвечающих за полезные признаки, является естественным развитием работ по селекции животных и растений, расширивших возможности селекционеров в части управляемости процесса создания новых сортов и расширения его возможностей, в частности, передачи полезных признаков между нескрещивающимися видами. Методы создания ГМО Основные этапы создания ГМО: 1. Получение изолированного гена. 2. Введение гена в вектор для переноса в организм. 3. Перенос вектора с геном в модифицируемый организм. 4. Преобразование клеток организма. 5. Отбор генетически модифицированных организмов и устранение тех, которые не были успешно модифицированы. Процесс синтеза генов в настоящее время разработан очень хорошо и даже в значительной степени автоматизирован. Существуют специальные аппараты, снабжённые ЭВМ, в памяти которых закладывают программы синтеза различных нуклеотидных последовательностей. Чтобы встроить ген в вектор, используют ферменты — рестриктазы и лигазы. С помощью рестриктаз ген и вектор можно разрезать на кусочки. С помощью лигаз такие кусочки можно «склеивать», соединять в иной комбинации, конструируя новый ген или заключая его в вектор. Если модификации подвергаются одноклеточные организмы или культуры клеток многоклеточных, то на этом этапе начинается клонирование, то есть отбор тех организмов и их потомков (клонов), которые подверглись модификации. Когда же поставлена задача получить многоклеточные организмы, то клетки с изменённым генотипом используют для вегетативного размножения растений Виды ГМО На сегодня в мире существует несколько десятков линий ГМ-культур: сои, картофеля, кукурузы, сахарной свеклы, риса, томатов, рапса, пшеницы, дыни, цикория, папайи, кабачков, хлопка, льна и люцерны. Массово выращиваются ГМ-соя, которая в США уже вытеснила обычную сою, кукуруза, рапс и хлопок. Посевы трансгенных растений постоянно увеличиваются. В 1996 году в мире под посевами трансгенных сортов растений было занято 1,7 млн. га, в 2006 году – 102 млн. га. В 2006 году ГМ-культуры выращивали в 22 странах мира, среди которых Аргентина, Австралия, Канада, Китай, Германия, Колумбия, Индия, Индонезия, Мексика, Южная Африка, Испания, США. Основные мировые производители продукции, содержащую ГМО – США (68%), Аргентина (11,8%), Канада (6%), Китай (3%). 3 Трансмиссивные и природно-очаговые заболевания Паразитарные болезни (паразитозы) в зависимости от природы возбудителя подразделяются на несколько групп. Заболевания, возбудители которых передаются только от животных к животным, называются зоонозы (чума кур и свиней). Заболевания, возбудители которых передаются только от человека к человеку, называются антропонозы (корь, дифтерия). Заболевания, возбудители которых передаются от одного организма к другому посредством кровососущих переносчиков (насекомые, клещи), называются трансмиссивными (малярия, таежный энцефалит). Они подразделяются на: 1) облигатно-трансмиссивные, возбудители которых передаются через специфических переносчиков (малярия — комарами рода Anopheles, таежный энцефалит— таежными клещами); 2) факультативно-трансмиссивные, возбудители которых могут передаваться как через переносчиков, так и другими путями (заражение туляремией и сибирской язвой возможно через многочисленных переносчиков и при разделке туш больных животных). Любая трансмиссивная болезнь представляет собой трехчленную систему, которая включает паразита — возбудителя, позвоночного — хозяина, и членистоногого — переносчика (клещи, насекомые). Переносчики возбудителей трансмиссивных болезней могут быть специфические и механические. В организме специфического переносчика возбудитель проходит часть жизненного цикла (чумная палочка размножается в пищеварительном тракте блохи; малярийные плазмодии проходят половой цикл развития у комаров рода Anopheles). Возбудители болезней у механических переносчиков (мухи, тараканы) находятся на покровах тела, на конечностях и частях ротового аппарата. Входные ворота возбудителя — всегда ротовой аппарат специфического переносчика. Выход возбудителя из переносчика может происходить через анальное отверстие или через ротовой аппарат. В первом случае происходит транзит возбудителя через кишечник (риккетсии вшивого сыпного тифа). Заражение хозяина происходит при втирании экскрементов переносчика в кожу при расчесах места укуса. Такой способ заражения называется контаминацией. Если возбудитель проходит через полость тела переносчика и накапливается в слюнных железах (спорозоиты малярийных плазмодиев), то заражение хозяина происходит через ротовой аппарат при кровососании. Способ заражения называется инокуляцией. Ворота выхода возбудителя могут отсутствовать. В таком случае возбудитель накапливается в полости тела переносчика. Заражение хозяина происходит при раздавливании переносчика и втирании гемолимфы с возбудителем в кожу при расчесах — разновидность контаминации (передача спирохет возвратного тифа вшами). В первом и втором случаях переносчик может передавать возбудителей многократно, в третьем — только однократно, так как передача возбудителя связана с гибелью переносчика. Для многих переносчиков характерна трансовариалъная (через яйца) передача возбудителей трансмиссивных заболеваний. Если самка таежного клеща содержит вирус энцефалита, то при половом размножении она передаст его последующим поколениям. В 1940 г. академик Е. Н. Павловский разработал учение о природной очаговости трансмиссивных болезней. Оно возникло на стыке экологии, паразитологии, эпидемиологии и ландшафтной географии. Представления о природной очаговости болезней (роль диких животных в передаче чумы) впервые были изложены в работах Д. К. Заболотного. В основе концепции природной очаговости болезней лежат экологические принципы. Природно-очаговыми называются болезни, связанные с комплексом природных условий. Они существуют в определенных биогеоценозах независимо от человека и для их поддержания важное значение имеют трофические связи. Е. Н. Павловский дал следующее определение природно-очаговых болезней: «Природная очаговость трансмиссивных болезней — это явление, когда возбудитель, специфический его переносчик и животные-резервуары возбудителя в течение смены своих поколений неограниченно долгое время существуют в природных условиях вне зависимости от человека, как по ходу своей уже прошедшей эволюции, так и в настоящий ее период». Природный очаг — это наименьшая территория одного или нескольких ландшафтов, где осуществляется циркуляция без заноса его извне неопределенно долгий срок. Компоненты природного очага заболевания: 1) возбудитель заболевания; 2) восприимчивые к данному возбудителю организмы; 3) переносчики возбудителя; 4) определенные условия среды (биотоп) Например: схема природного очага чумы Блоха- грызуны человек грызуны Восприимчивость - это видовое свойство, определяющее способность особей данного вида стать средой обитания для паразита-возбудителя и отвечать на его внедрение специфическими реакциями. В очаге происходит циркуляция возбудителя от больных животных (доноров возбудителя) через переносчика к здоровым (реципиентам), которые в дальнейшем становятся донорами возбудителя. Переносчиками являются кровососущие членистоногие, а донорами и реципиентами могут быть грызуны и птицы. Если человек попадает в природный очаг заболевания, то он становится сначала реципиентом, а затем и донором возбудителя. Природные очаги существуют длительное время, но эпидемиологическое значение они приобретают тогда, когда в них попадает и заражается человек (дикие животные, (дикие животные) чаще грызуны) Классификация природных очагов: По происхождению выделяют очаги: 1) природные (клещевой энцефалит); 2) синантропные— существуют в населенном пункте, где циркуляция возбудителя осуществляется за счет синантропных животных (чесотка); 3) антропургические— возникают в результате преобразования природной среды человеком (описторхоз в местах искусственно созданных водоемов); 4) смешанные (трихинеллез). Ареал природных очагов определяется ареалом естественных хозяев возбудителя и ареалом переносчика. Попротяженности (площади) очаги могут быть: 1) узко ограниченными(нора грызуна, гнездо птицы —очаг клещевого возвратного тифа); 2) диффузными(тайга — очаг таежного энцефалита); 3) сопряженными,если в очаге циркулируют возбудители нескольких трансмиссивных болезней (туляремии и чумы). Результатом заражения реципиента в природном очаге может быть его гибель (в случае высокой вирулентности возбудителя), болезнь с последующим выздоровлением или вакцинация (образование иммунных защитных телбез выраженных клинических признаков болезни - при слабой вирулентности возбудителя). На исход заражения реципиента в очаге также влияют следующие факторы: 1) патогенность возбудителя для данного реципиента; 2) «агрессивность» переносчика (частота кровососания); 3) доза возбудителя, вводимого в организм реципиента; 4) степень выраженности неспецифических и специфических иммунных реакций реципиента. Некоторые природно-очаговые заболевания характеризуются эндемизмом, т.е. встречаемостью на строго ограниченных территориях. Это связано с тем, что возбудители соответствующих заболеваний, их промежуточные хозяева, животные-резервуары или переносчики встречаются только в определенных биогеоценозах. Так, только в отдельных районах Японии расселены четыре вида легочных сосальщиков из p. Paragonimus. Расселению их препятствует узкая специфичность в отношении промежуточных хозяев, которые обитают только в некоторых водоемах Японии, а природным резервуаром являются такие эндемичные виды животных, как японская луговая мышь или японская куница. Вирусы некоторых форм геморрагической лихорадки встречаются только в определенных зонах Восточной Африки, потому что здесь расположен ареал их специфических переносчиков — клещей из р. Аmbliomma. Хорошо известными являются природно-очаговые заболевания, вызываемые лентецами p. Diphyllobothrium, распространенными в некоторых районах Сибири. Так, на оз. Байкал известно несколько видов ленточных червей, которые обычно паразитируют у чаек, заражающихся при проглатывании инвазированной рыбы, например байкальского омуля. Рыбы заражаются при поедании рачков, а те в свою очередь становятся промежуточными хозяевами гельминтов от фекалий птиц, попадающих в воду. Съев инвазированную рыбу, заболеть дифиллоботриозом может и человек. Существуют природно-очаговые заболевания, имеющие более широкий ареал. Так, в бассейне р. Оби и Иртыша, а также в некоторых других зонах Сибири и Восточной Европы распространено заболевание описторхоз, встречающееся у медведей, выдр, кошек, волков, лис, а также у человека. Возбудителем этого заболевания является кошачий сосальщик, расселение которого ограничивается, вероятно, комплексом природно-климатических факторов. Об этом свидетельствует то, что ближайший родственник этого паразита, сосальщик виверры, встречается только в Юго-Восточной Азии и не выходит за пределы этого региона, являясь, таким образом, компонентом восточно-азиатских тропических биогеоценозов. Небольшое количество природно-очаговых заболеваний встречается практически повсеместно. Это такие заболевания, возбудители которых, как правило, не связаны в цикле своего развития с внешней средой и поражают самых разнообразных хозяев. К заболеваниям такого рода относятся, например, токсоплазмоз и трихинеллез. Этими природно-очаговыми болезнями человек может заразиться в любой природно-климатической зоне и в любой экологической системе. Абсолютное же большинство природно-очаговых болезней поражает человека только в случае попадания его в соответствующий очаг (на охоте, рыбной ловле, в туристических походах, в геологических партиях и т.д.) при условиях его восприимчивости к ним. Так, таежным энцефалитом человек заражается при укусе инфицированным клещом, а описторхозом — съев недостаточно термически обработанную рыбу с личинками кошачьего сосальщика. Профилактика природно-очаговых заболеваний представляет особые сложности. В связи с тем что в циркуляцию возбудителя бывает включено большое количество хозяев, а часто и переносчиков, разрушение целых биогеоценотических комплексов, возникших в результате эволюционного процесса, экологически неразумно, вредно и даже технически невозможно. Лишь в тех случаях, если очаги являются небольшими и хорошо изученными, возможно комплексное преобразование таких биогеоценозов в направлении, исключающем циркуляцию возбудителя. Так, рекультивация опустыненных ландшафтов с созданием на их месте орошаемых садоводческих хозяйств, проводящаяся на фоне борьбы с пустынными грызунами и москитами, может резко снизить заболеваемость населения лейшманиозами. В большинстве же случаев природно-очаговых болезней профилактика их должна быть направлена в первую очередь на индивидуальную защиту (предотвращение от укусов кровососущими членистоногими, термическая обработка пищевых продуктов и т.д.) в соответствии с путями циркуляции в природе конкретных возбудителей. билет 44 1 К фундаментальным свойствам живых систем относятся: 1. ОСОБЕННОСТИ ХИМИЧЕСКОГО СОСТАВА - в состав живых организмов входят те же химические элементы, что и в объекты неживой природы, однако, соотношение различных элементов в живом и неживом неодинаково. Элементарный состав неживой природы наряду с кислородом представлен, в основном, кремнием, железом, магнием, алюминием. В живых организмах 98% их массы приходится на: водород, кислород, углерод и азот; 2. ОБМЕН ВЕЩЕСТВ - все живые организмы способны к обмену веществ с окружающей средой, поглощая из нее вещества, необходимые для питания, и выделяя наружу продукты жизнедеятельности. В круговороте органических веществ самыми существенными являются процессы синтеза и распада живыми веществами поглощенных из окружающей среды различных веществ. В результате целого ряда сложных химических превращений вещества из окружающей Среды уподобляются веществам живых организмов и из них строятся тела живых организмов. Эти процессы называются АССИМИЛЯЦИЕЙ(пластический обмен или АНАБОЛИЗМ). С другой стороны происходят процессы ДИССИМИЛЯЦИИ(КАТАБОЛИЗМ), в результате которых сложные органические соединения распадаются на простые и выделяется энергия, необходимая для реакций биосинтеза. Обмен веществ обеспечивает постоянство химического состава и строения всех частей организма и, как следствие, постоянство их функционирования в непрерывно меняющихся условиях окружающей среды. 3. САМОВОСПРОИЗВЕДЕНИЕ (РЕПРОДУКЦИЯ) - способность к размножению, то есть воспроизведению нового поколения особей того же вида. В основе самовоспроизведения лежат реакции матричного синтеза, то есть образование новых молекул и структур на основе информации, заложенной в последовательности нуклеотидов ДНК. 4. НАСЛЕДСТВЕННОСТЬ - заключается в способности организмов передавать свои признаки, свойства и особенности развития из поколения в поколение. Она обусловлена стабильностью, то есть постоянством строения молекул ДНК. 5. ИЗМЕНЧИВОСТЬ - способность организмов приобретать новые признаки и свойства. В основе ее лежит изменение биологических матриц. Если бы репродукция матриц - молекул ДНК - всегда происходила с абсолютной точностью, то приспособление видов к меняющимся условиям среды оказалось бы невозможным. Изменчивость создает разнообразный исходный материал для есте-ственного отбора. 6. РОСТ и РАЗВИТИЕ. Под развитием понимают необратимое направленное закономерное изменение объектов живой и неживой природы. Развитие живой формы существования материи представлено инди-видуальным развитием или ОНТОГЕНЕЗОМ и историческим развитием или ФИЛОГЕНЕЗОМ (эволюция). 7. РАЗДРАЖИМОСТЬ - свойство избирательно реагировать на изменение внешней и внутренней среды. Реакция многоклеточных животных на раздражение осуществляется посредством нервной системы и называется РЕФЛЕКСОМ. Организмы, не имеющие нервной системы лишены рефлексов. Их реакции принято называть ТАКСИСАМИ или ТРОПИЗМАМИ. Например: фототаксис - движение по отношению к источнику света; хемотаксис - перемещение организма по отношению к концентрации химических веществ. Под тропизмом понимают характер роста, свойственный растениям. Например: гелиотропизм - рост наземных частей растения по отношению к Солнцу. Геотропизм - рост подземных частей (корней) в направлении к центру Земли. 8. ДИСКРЕТНОСТЬ. Жизнь на Земле проявляется в виде дискретных форм. Это означает, что отдельный организм или иная биологическая система (вид, биоценоз и др.) состоит из отдельных изолированных, но тесно связанных и взаимодействующих между собой частей, образующих структурно-функциональное единство. Например, любой вид организмов представлен отдельными особями. Дискретность строения организма - основа его структурной упорядоченности, она создает возможность постоянного самообновления его путем замены “износившихся” структурных элементов. 9. САМОРЕГУЛЯЦИЯ (АВТОРЕГУЛЯЦИЯ) - способность живых организмов, обитающих в непрерывно меняющихся условиях среды, поддерживать постоянство своего химического состава и интенсивность течения физиологических процессов. 10. РИТМИЧНОСТЬ (присуща как живой, так и неживой природе) -обусловлена различными космическими и планетарными причинами. РИТМ - это повторение одного и того же события или воспроизведение одного и того же состояния через равные промежутки времени. Ритмичность направлена на согласование функций живых организмов с окружающей средой, то есть приспособление к изменяющимся условиям существования. 11. ЭНЕРГОЗАВИСИМОСТЬ. Живые тела - “открытые” для поступления энергии системы. Под “открытыми” системами понимают системы, в которых непрерывно происходит поглощение и удаление веществ, а также обмен энергией со средой. Таким образом, “ЖИВЫЕ ТЕЛА, СУЩЕСТВУЮЩИЕ НА ЗЕМЛЕ, ПРЕДСТАВЛЯЮТ СОБОЙ ОТКРЫТЫЕ САМОРЕГУЛИРУЮЩИЕСЯ И САМОВОСПРОИЗВОДЯЩИЕСЯ СИСТЕМЫ, ПОСТРОЕННЫЕ ИЗ БИОПОЛИМЕРОВ И НУКЛЕИНОВЫХ КИСЛОТ” (М.В.Волькенштейн). 2.Обратная транскрипция, вирусная трансдукция и их значение для генетической инженерии. Понятие о векторных молекулах. Обратная транскрипция — это процесс образования двуцепочечной ДНК на матрице одноцепочечной РНК. Данный процесс называется обратной транскрипцией, так как передача генетической информации при этом происходит в «обратном», относительно транскрипции, направлении. Идея обратной транскрипции вначале была очень непопулярна, так как противоречила центральной догме молекулярной биологии, которая предполагала, что ДНК транскрибируется в РНК и далее транслируется в белки. Встречается у ретровирусов, например, ВИЧ и в случае ретротранспозонов. Трансдукция— процесс переноса бактериальной ДНК из одной клетки в другую бактериофагом. Общая трансдукция используется в генетике бактерий для картирования генома и конструирования штаммов. К трансдукции способны как умеренные фаги, так и вирулентные, последние, однако, уничтожают популяцию бактерий, поэтому трансдукция с их помощью не имеет большого значения ни в природе, ни при проведении исследований. Векторная молекула ДНК — это молекула ДНК, которая выступает в роли носителя. Молекулу-носитель должен отличать ряд особенностей: - Способность к автономной репликации в клетке хозяина (чаще бактериальной или дрожжевой) - Наличие селективного маркера - Наличие удобных сайтов рестрикции В роли векторов чаще всего выступают бактериальные плазмиды. 3 В определении половой принадлежности организма Роль наследственных и средовых факторов Соотносительная роль наследственной программы и факторов среды в формировании фенотипа особи может быть прослежена на примере развития признаков половой принадлежности организма. Пол организма представляет собой важную фенотипическую характеристику, которая проявляется в совокупности свойств, обеспечивающих воспроизведение потомства и передачу ему наследственной информации. В зависимости от значимости этих свойств различают первичные и вторичные половые признаки. Под первичными половыми признаками понимают морфофизиологические особенности организма, обеспечивающие образование половых клеток — гамет, сближение и соединение их в процессе оплодотворения. Это наружные и внутренние органы размножения. Вторичными половыми признаками называют отличительные особенности того или другого пола, не связанные непосредственно с гаметогенезом, спариванием и оплодотворением, но играющие важную роль в половом размножении (обнаружение, и привлечение партнера). Их развитие контролируется гормонами, синтезируемыми первичными половыми органами. Важным доказательством в пользу наследственной детерминированности половой принадлежности организмов является наблюдаемое у большинства видов соотношение по полу 1:1 (табл. 6.1). Таблица 6.1.Соотношение по полу у разных видов 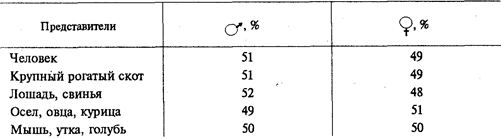 Такое соотношение может быть обусловлено образованием двух видов гамет представителями одного пола (гетерогаметный пол) и одного вида гамет — особями другого пола (гомогаметный пол). Это соответствует различиям в кариотипах организмов разных полов одного и того же вида, проявляющимся в половых хромосомах. У гомогаметного пола, имеющего одинаковые половые хромосомыXX,все гаметы несут гаплоидный набор аутосом плюс Х-хромосому. У гетерогаметного пола в кариотипе кроме аутосом содержатся две разные или только одна половая хромосома (XY или ХО). Его представители образуют два вида гамет, различающиеся по гетерохромосомам: Х и Y или Х и 0. У разных видов организмов хромосомный механизм определения пола реализуется по-разному. У человека и других млекопитающих, а также у дрозофилы гомогаметным является женский пол (XX), а гетерогаметным — мужской (XY). У некоторых насекомых (клопы рода Protenor) гетерогаметный мужской пол имеет лишь одну Х-хромосому (ХО). У птиц и некоторых насекомых женский пол является гетерогаметным (XY), а мужской — гомогаметным (XX). У некоторых бабочек гетерогаметный женский пол имеет одну Х-хромосому (ХО). В большинстве выше описанных случаев пол вновь образующегося организма определяется сочетанием половых хромосом, возникающим в зиготе при оплодотворении (табл. 6.2). Таблица 6.2.Хромосомный механизм определения полов 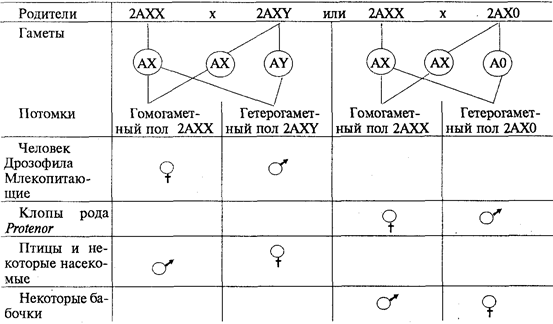 Таким образом, хромосомный механизм определения половой принадлежности организмов обеспечивает равновероятность встречаемости представителей обоих полов. Это имеет большой биологический смысл, так как обусловливает максимальную вероятность встречи самки и самца, потомки получают более разнообразную наследственную информацию, поддерживается оптимальная численность особей в популяции. Исследования, проведенные на дрозофиле, обнаружили незначительную роль Y-хромосомы в определении пола. Особи с кариотипом ХО внешне являются типичными самцами, но стерильными, а особи с кариотипом XXY — плодовитые самки. Y-хромосома у дрозофилы по размеру близка к Х-хромосоме, однако она генетически инертна, так как состоит в основном из гетерохроматина. В связи с этим первичное определение пола у дрозофилы обусловлено не столько сочетанием в зиготе Х- и Y-хромосом, сколько соотношением числа Х-хромосом и наборов аутосом (половой индекс). У нормальных самок половой индекс равен 1 (2Х:2А), у нормальных самцов — 0,5 (XY:2A). Увеличение полового индекса более 1 при увеличении дозы Х-хромосомы (ЗХ:2А) приводит к развитию сверхсамок, а величина этого показателя ниже 0,5 (XY:3A) обеспечивает появление самцов. При значениях полового индекса более 0,5, но менее 1 (2Х:ЗА) развиваются интерсексы. О значении дозы Х-хромосом в определении пола у дрозофилы свидетельствует развитие латерального гинандроморфизма у мух при утрате Х-хромосомы одной из клеток на стадии первого деления зиготы. В результате половина тела, развивающаяся из клетки с нормальным кариотипом (2АХХ), несет признаки самки, а другая половина, клетки которой лишены одной Х-хромосомы (2АХО), имеет признаки самца (рис. 6.2). 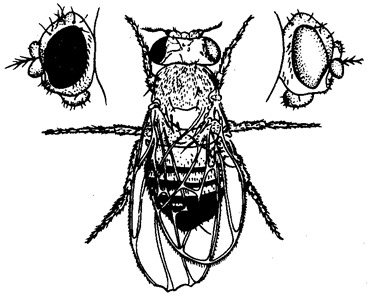 Рис. 6.2. Латеральный гинадроморфизм у Drosophila melanogaster, гетерозиготной по гену окраски глаз, находящемуся в Х-хромосоме Левая половина мухи, клетки которой несут 2Х, имеет признаки самки и красный глаз; правая половина мухи, клетки которой несут одну Х-хромосому, имеет признаки самца и белый глаз Обнаружение зависимости половой принадлежности развивающегося организма от дозы Х-хромосом у дрозофилы и некоторых других насекомых привело американского генетика С. Бриджеса (1922) к формулировке гипотезы генного баланса, в соответствии с которой организм изначально бисексуален, т.е. несет в себе задатки и мужского, и женского полов. Развитие признаков одного из них в ходе онтогенеза определяется балансом женских и мужских генов — детерминаторов пола. У дрозофилы эти гены сосредоточены не только в половой Х-хромосоме, но и в аутосомах. Поэтому пол организма у них зависит от соотношения этих хромосом. У плодовой мухи Y-хромосома, будучи генетически инертной, в определении признаков пола не участвует. У человека Y-хромосома играет важную роль в детерминации пола. Она содержит определенное количество генов, часть из которых гомологична генам Х-хромосомы, а часть не имеет в ней гомологов и наследуется только по мужской линии. Некоторые из этих генов непосредственно связаны с детерминацией мужского пола. Поэтому у человека присутствие Y-хромосомы в кариотипе независимо от количества Х-хромосом (2AXXY, 2AXXXY) обеспечивает развитие мужского пола. Особи с кариотипом 2АХО являются женщинами, несмотря на уменьшение дозы Х-хромосомы. Однако значение баланса генов в определении половой принадлежности организма подтверждается тем, что особи с кариотипами 2АХО, 2АХХХ, 2AXXY, 2AXXXY и т.д. отличаются наличием пороков развития и часто стерильны. Как было отмечено, у большинства видов животных пол организма определяется сочетанием или количеством половых хромосом в диплоидном наборе аутосом. Однако у некоторых насекомых (пчелы, муравьи, осы) представители разных полов отличаются числом наборов хромосом. Самки у них развиваются из оплодотворенных яиц и поэтому диплоидны, самцы — из неоплодотворенных яиц и первично гаплоидны. В такой ситуации соотношение по полу зависит от того, происходит оплодотворение или нет. Половой состав популяции может быть различным в разные периоды ее жизни. Зависимость половой принадлежности организмов от сочетания или количества половых хромосом в его кариотипе свидетельствует о том, что гены, определяющие пол, располагаются главным образом в этих хромосомах. Однако известен целый ряд примеров, указывающих на то, что в развитии признаков пола принимают участие также гены, локализованные в аутосомах. В частности, в геноме дрозофилы присутствует ген, называемый трансформатором. Рецессивный аллель этого гена в гомозиготном состоянии tt превращает женские зиготы 2Х в фенотипических самцов, которые оказываются стерильными. Самки1 XXt+t+ или XXt+t плодовиты, особи с кариотипом XYtt — нормальные самцы, организмы с кариотипом XXtt — превращенные стерильные самцы. Другим примером влияния аутосомных генов на развитие признаков пола может служить существование у кукурузы генов, мутантные аллели которых превращают ее из однодомного растения в двудомное, препятствуя развитию органов либо мужского, либо женского пола. Рецессивный аллель гена sk(silkless) в гомозиготном состоянии вызывает недоразвитие семяпочек и превращает растение в мужское. Мутантный аллель другого гена ts (tassel seed) в гомозиготном состоянии вызывает развитие семяпочек и семян в метелке, а пыльники не развиваются. У человека развитие организма по мужскому типу обеспечивается не только геном, расположенным в Y-хромосоме и определяющим способность к синтезу мужского полового гормона — тестостерона, но и Х-сцепленным геном, контролирующим синтез белка-рецептора этого гормона. Мутация упомянутого Х-сцепленного гена приводит к развитию синдрома тестикулярной феминизации (см. разд. 3.6.5.2). Приведенные примеры доказывают, что пол организмов как признак развивается на основе полученной от родителей наследственной информации и находится под контролем группы взаимодействующих генов, расположенных как в половых хромосомах, так и в аутосомах. билет 45 1 1. Уровни организации жизни Различают следующие уровни организации органического мира: В микросистемы входят: а) молекулярно-генетический уровень б) субклеточный в) клеточный уровни. Биологически мезосистемы: а) тканевый уровень б) органный уровень в) организменный Биологические макросистемы: а) популяционно-видовой б) биоценотический в) биосферный. Молекулярный уровень организации - это уровень функционирования биологических макромолекул - биополимеров: нуклеиновых кислот, белков, полисахаридов, липидов, стероидов. С этого уровня начинаются важнейшие процессы жизнедеятельности: обмен веществ, превращение энергии, передача наследственной информации. Этот уровень изучают: биохимия, молекулярная генетика, молекулярная биология, генетика, биофизика. Клеточный уровень - это уровень клеток (клеток бактерий, цианобактерий, одноклеточных животных и водорослей, одноклеточных грибов, клеток многоклеточных организмов). Клетка - это структурная единица живого, функциональная единица, единица развития. Этот уровень изучают цитология, цитохимия, цитогенетика, микробиология. Тканевый уровень организации - это уровень, на котором изучается строение и функционирование тканей. Исследуется этот уровень гистологией и гистохимией. Органный уровень организации - это уровень органов многоклеточных организмов. Изучают этот уровень анатомия, физиология, эмбриология. Организменный уровень организации - это уровень одноклеточных, колониальных и многоклеточных организмов. Специфика организменного уровня в том, что на этом уровне происходит декодирование и реализация генетической информации, формирование признаков, присущих особям данного вида. Этот уровень изучается морфологией (анатомией и эмбриологией), физиологией, генетикой, палеонтологией. Популяционно-видовой уровень - это уровень совокупностей особей - популяций и видов. Этот уровень изучается систематикой, таксономией, экологией, биогеографией, генетикой популяций. На этом уровне изучаются генетические и экологические особенности популяций, элементарные эволюционные факторы и их влияние на генофонд (микроэволюция), проблема сохранения видов. Экосистемный уровень организации - это уровень микроэкосистем, мезоэкосистем, макроэкосистем. На этом уровне изучаются типы питания, типы взаимоотношений организмов и популяций в экосистеме, численность популяций, динамика численности популяций, плотность популяций, продуктивность экосистем, сукцессии. Этот уровень изучает экология. Выделяют также биосферный уровень организации живой материи. Биосфера - это гигантская экосистема, занимающая часть географической оболочки Земли. Это мегаэкосистема. В биосфере происходит круговорот веществ и химических элементов, а также превращение солнечной энергии. 2 Структурные элементы ядра бывают четко выражены только в определенный период клеточного цикла в интерфазе. В период деления клетки (в период митоза или мейоза) одни структурные элементы исчезают, другие существенно преобразуются. Классификация структурных элементов интерфазного ядра: хроматин; ядрышко; кариоплазма; кариолемма. Хроматин представляет собой вещество, хорошо воспринимающее краситель (хромос), откуда и произошло его название. Хроматин состоит из хроматиновых фибрилл, толщиной 20-25 нм, которые могут располагаться в ядре рыхло или компактно. На этом основании различают два вида хроматина: · эухроматин - рыхлый или деконденсированный хроматин, слабо окрашивается основными красителями; · гетерохроматин - компактный или конденсированный хроматин, хорошо окрашивается этими же красителями. При подготовке клетки к делению в ядре происходит спирализация хроматиновых фибрилл и превращение хроматина в хромосомы. После деления в ядрах дочерних клеток происходит деспирализация хроматиновых фибрилл и хромосомы снова преобразуются в хроматин. Следовательно, хроматин и хромосомы представляют собой различные фазы одного и того же вещества. По химическому строениюхроматин состоит из: дезоксирибонуклеиновой кислоты (ДНК) 40 %; белков около 60 %; рибонуклеиновой кислоты (РНК) 1 %. Ядерные белки представлены формами: щелочными или гистоновыми белками 80-85 %; кислыми белками 15-20 %. Гистоновые белки связаны с ДНК и образуют полимерные цепи дезоксирибонуклеопротеида (ДНП), которые и представляют собой хроматиновые фибриллы, отчетливо видимые при электронной микроскопии. На определенных участках хроматиновых фибрилл осуществляется транскрипция с ДНК различных РНК, с помощью которых осуществляется затем синтез белковых молекул. Процессы транскрипции в ядре осуществляются только на свободных хромосомных фибриллах, то есть в эухроматине. В конденсированном хроматине эти процессы не осуществляются и потому гетерохроматин является неактивным хроматином. Соотношение эухроматина и гетерохроматина в ядре является показателем активности синтетических процессов в данной клетке. На хроматиновых фибриллах в S-периоде интерфазы осуществляется также процессы редупликации ДНК. Эти процессы происходят как в эухроматине, так и в гетерохроматине, но в гетерохроматине они протекают значительно позже. Ядрышко - сферическое образование (1-5 мкм в диаметре) хорошо воспринимающее основные красители и располагающееся среди хроматина. В одном ядре может содержаться от 1 до 4-х и даже более ядрышек. В молодых и часто делящихся клетках размер ядрышек и их количество увеличены. Ядрышко не является самостоятельной структурой. Оно формируется только в интерфазе в определенных участках некоторых хромосом - ядрышковых организаторах, в которых содержатся гены, кодирующие молекулу рибосомальной РНК. В области ядрышкового анализатора осуществляется транскрипция с ДНК рибосомальной РНК. В ядрышке происходит соединение рибосомальной РНК с белком и образование субъединиц рибосом. Микроскопически в ядрышке различают: · фибриллярный компонент - локализуется в центральной части ядрышка и представляет собой нити рибонуклеопротеида (РНП); · гранулярный компонент - локализуется в периферической части ядрышка и представляет скопление субъединиц рибосом. В профазе митоза, когда происходит спирализация хроматиновых фибрилл и образование хромосом, процессы транскрипции РНК и синтеза субъединиц рибосом прекращаются и ядрышко исчезает. По окончании митоза в ядрах вновь образованных клеток происходит деконденсация хромосом и появляется ядрышко. Кариоплазма (нуклеоплазма) или ядерный сок состоит из воды, белков и белковых комплексов (нуклеопротеидов, гликопротеидов), аминокислот, нуклеотидов, сахаров. Под световым микроскопом кариоплазма бесструктурна, но при электронной микроскопии в ней определяются гранулы (15 нм), состоящие из рибонуклеопротеидов. Белки кариоплазмы являются в основном белками-ферментами, в том числе ферментами гликолиза, осуществляющих расщепление углеводов и образование АТФ. Негистоновые (кислые) белки образуют в ядре структурную сеть (ядерный белковый матрикс), которая вместе с ядерной оболочкой принимает участие в создание внутреннего порядка, прежде всего в определенной локализации хроматина. При участии кариоплазмы осуществляется обмен веществ в ядре, взаимодействие ядра и цитоплазмы. Кариолемма (нуклеолемма) - ядерная оболочка отделяет содержимое ядра от цитоплазмы (барьерная функция), в то же время обеспечивает регулируемый обмен веществ между ядром и цитоплазмой. Ядерная оболочка принимает участие в фиксации хроматина. Кариолемма состоит из двух билипидных мембран - внешней и внутренней ядерной мембраны, разделенных перинуклеарным пространством, шириной от 25 до 100 нм. В кариолемме имеются поры, диаметром 80-90 нм. В области пор внешняя и внутренняя ядерные мембраны переходят друг в друга, а перинуклеарное пространство оказывается замкнутым. Просвет поры закрыт особым структурным образованием - комплексом поры, который состоит из фибриллярного и гранулярного компонента. Гранулярный компонент представлен белковыми гранулами диаметром 25 нм, располагающимися по краю поры в три ряда. От каждой гранулы отходят фибриллы и соединяются в центральной грануле, располагающейся в центре поры. Комплекс поры играет роль диафрагмы, регулирующей ее проницаемость. Размеры пор стабильны для данного типа клеток, но число пор может изменяться в процессе дифференцировки клетки. В ядрах сперматозоидов ядерные поры отсутствуют. На наружной ядерной мембране могут локализоваться прикрепленные рибосомы. Кроме того, наружная ядерная мембрана может продолжаться в канальцы эндоплазматической сети. 3 |