Белки общие свойства. 1. Белки структура и функции
 Скачать 2.19 Mb. Скачать 2.19 Mb.
|

3. Классификация белковВ основе классификации лежат: 1. Физико-химические факторы: кислые, основные и нейтральные белки, а также гидрофильные, гидрофобные и амфифильные. 2. Функциональные свойства В основе лежат биологические свойства белков – каталитические, защитные, транспортные, регуляторные, механические, сократительные и др. 3. Структурные признаки: а) простые белки – протеины, представлены только полипептидной цепью; б) сложные белки – протеиды – в основе классификации лежит характер простетической группы (небелковая часть): – гликопротеины (небелковая часть представлена углеводами); – липопротеины (небелковая часть представлена липидами); – нуклеопротеины (небелковая часть представлена нуклеиновыми кислотами); – витаминпротеины (небелковая часть представлена витаминами); – хромопротеины (небелковая часть представлена окрашенными соединениями, например, гем гемоглобина); – металлопротеины (небелковая часть представлена металлами); – фосфопротеины (небелковая часть представлена остатками фосфорной кислоты, например, в казеине молока). 3.1. Простые белкиК этой группе белков относят гистоны, протамины, альбумины, глобулины, проламины, глютелины и протеиноиды (α-кератин, фиброин, коллаген, эластин). Гистоны – тканевые белки многоклеточных организмов, локализованы в ядре клеток. Представляют собой низкомолекулярные (Мм 11000–24000) белки с третичной структурой, обладают выраженными основными свойствами (рI 9,5–12), т.к. содержат большое количество аргинина и лизина. Гистоны прочно связаны с ДНК хроматина (связь ионная). Выделяют пять фракций гистонов (методом электрофореза в полиакриламидном геле, ПААГ). Разделение на фракции зависит от соотношения в них аргинина и лизина. Основные функции гистонов – структурная и регуляторная: – структурная – гистоны участвуют в стабилизации пространственной структуры ДНК, а следовательно хроматина, хромосом и нуклеосом; – регуляторная – заключается в способности блокировать передачу генетической информации от ДНК к РНК (транскрипцию) (рис. 14). 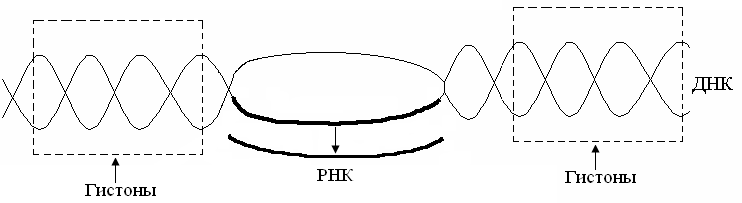 Рис. 14. Транскрипция ДНК Участки ДНК, связанные с гистонами, не могут подвергаться транскрипции. Там, где ДНК свободна от гистонов, двойная спираль расплетается и освободившийся участок ДНК подвергается транскрипции. Протамины – самые низкомолекулярные белки (Мм 4000–12000). Резко выражен основный характер, так как содержат до 80% аргинина. Протамины связывают ДНК в хроматине спермиев и делают ее компактной. Выполняют главным образом структурную функцию, поэтому и присутствуют в клетках не способных к делению. Альбумины и глобулины – содержатся в плазме крови, в клетках тканей, молоке, клетках растений, зерне и др. Они разнообразны по структуре, физико-химическим свойствам и биологической роли. Альбумины – белки с Мм 15000–70000, следовательно, это мелкодисперсные белки. Имеют выраженный кислый характер (рI 4,7) так как содержат большое количество аспарагиновой и глутаминовой кислот. Хорошо растворимы в воде. Биологические функции альбуминов: 1. Трофическая (питательная) – альбумин куриного яйца (овальбумин), молока (лактальбумин), зерновых культур и другие. 2. Транспортная – альбумины обладают высокой адсорбционной способностью (адсорбируют полярные и неполярные молекулы). Например, альбумины крови транспортируют жирные кислоты от жирового депо к другим органам и тканям. 3. Обезвреживающая – альбумины связывают токсичные соединения – алкалоиды, тяжелые металлы, билирубин. 4. Альбумины обеспечивают на 80% осмотическое давление крови от всех других сывороточных белков. Глобулины – это слабокислые или нейтральные белки (рI 6,0–7,3), Мм >100000. Они нерастворимы в воде, но растворяются в слабо-солевых растворах нейтральных солей (например, 0,9% NaCl), поэтому мало гидратированы. Глобулины являются неоднородными белками. При электрофорезе на хроматографической бумаге их можно разделить в зависимости от электрофоретической подвижности на 4 фракции: α1-, α2-, β-, γ-глобулины. Биологические функции глобулинов: 1. Транспортная – в крови глобулины транспортируют холестерин, фосфолипиды, триглицериды, гаптоглобин, церулоплазмин, трансферрин, ионы железа (Fe2+), меди (Cu2+), витамин В12. В молоке лактоглобулины и лактальбумины также выполняют транспортную функцию. 2. Гемостатическая – β-глобулины содержат протромбин (свертывающая система) 3. Защитная – фракция γ-глобулинов представлена смесью различных иммуноглобулинов. α1-глобулины транспортируют острофазовый белок α1-кислый гликопротеин. 4. Трофическая – глобулины составляют основную часть запасных белков семядолей бобовых культур (сои, гороха, фасоли, вики), алейроновых зерен масличных культур (подсолнечника, хлопчатника, льна, горчицы) и овощных культур. 5. Альбумины и глобулины зародыша и алейронового слоя выполняют каталитическую (ферментативную) функцию при прорастании зерна и в технологических процессах. Проламины – белки растительного происхождения, содержатся в клейковине семян злаковых растений. В их состав входит большое количество глутамина (14–43%), пролина (6,3–19,0%) и малое количество ионогенных групп (дикарбоновые аминокислоты почти полностью амидированы, RCONH2). Проламины отличаются низким содержанием лизина. Очень мало его в зеине кукурузы (0,2%), глиадине пшеницы и секалине ржи (0,6–0,7%). Высокий процент лизина (3,3%) наблюдается в авенине овса (перечислены названия проламинов различных злаковых культур). Проламины также бедны треонином, триптофаном, аргинином и гистидином. Зеин кукурузы, оризин риса, кафирин сорго отличаются высоким содержанием лейцина (16,9–18,6%). По содержанию метионина выделяют глиадин пшеницы – 1,2%, авенин овса – 3,7%. Небольшое количество указанных аминокислот в проламинах компенсируется относительно большим содержанием данной фракции в суммарном белке большинства злаков. Проламины нерастворимы в воде, солевых растворах, кислотах и щелочах. Они растворяются в 70%-ном спирте, что связано с наличием большого количества неполярной аминокислоты пролина. Проламины практически отсутствуют в бобовых и масличных культурах. Глютелины – растительные белки, растворимые в щелочах, что объясняется большим содержанием аргинина. Они занимают промежуточное положение между проламинами и глобулинами. Содержание лизина, аргинина, и гистидина в них больше чем в проламинах. Так, содержание лизина в глютелине пшеницы составляет 2,6%, в глютелине ржи – 2,3%, ячменя – 4,0%, овса – 5%. Глютелины ячменя, риса и овса отличаются от глютелина пшеницы (глютенина) более высоким уровнем лизина. Например, у риса 80% всего белка приходится на глютелины (оризенин), этим можно объяснить удовлетворительное содержание лизина (2,6–4,0%) в общем белке рисового зерна. Преобладающими фракциями овса являются глобулины и глютелины, содержащие 5–5,5% лизина, что также обеспечивает хорошую сбалансированность данной культуры по лизину. Протеиноиды – белки опорных тканей (костей, хрящей, связок, сухожилий, ногтей, волос, эпидермиса кожи). Это фибриллярные белки, имеют β-конфигурацию полипептидных цепей, молекулы которых образуют многомолекулярные нитевидные комплексы – фибриллы. Самые распространенные в природе: фиброин, коллаген, кератин, эластин. Они могут иметь третичную и четвертичную структуры. Протеиноиды нерастворимы в воде – только в специальных растворителях. Не перевариваются в желудочно-кишечном тракте большинства животных и человека. Однако, некоторые членистоногие приспособились к питанию фибриллярными белками кожи, перьев птиц, шерсти (например, моль). Характеристика представителей протеиноидов: Фиброин – белок шелка. Его β-структура состоит из антипараллельных полипептидных цепей, связанных между собой водородными связями. α-Кератин – составляет эпидермис кожи, волосы, ногти, рога. Полипептидные цепи этих белков имеют конформацию α-спирали. В волосе три такие цепи, скрученные в суперспираль, образуют протофибриллу. Несколько протофибрилл вновь образуют спиральный жгут – микрофибриллу. Жгут из микрофибрилл образует макрофибриллу. Многожильный канат одного волоса содержит сотни макрофибрилл. Молекулы α-кератина ориентированы параллельно и соединены дисульфидными связями (содержат много цистеина), что придает прочность структуре. Коллаген – самый распространенный белок в мире животных. Он составляет 1/3 от общего количества белка позвоночных. Более 80% всего коллагена организма находится в межклеточном веществе соединительной ткани кожи, костей, связок, сухожилий, хрящей. Эти ткани обладают малой растяжимостью и высокой прочностью. Полипептидная цепь коллагена содержит приблизительно 1000 аминокислот. Она построена из повторяющейся тройки аминокислот – гли-А-В, где А и В чаще всего представлены пролином или гидроксипролином. На долю глицина приходится 30%, пролина и гидроксипролина – 20%, аланина – 10%, остальных аминокислот – 40%. Коллаген единственный белок, в состав которого входит гидроксипролин. Полипептидные цепи коллагена при формировании вторичной и третичной структуры не могут давать типичные α-спирали, так как этому мешают антиспиральные аминокислоты – глицин, пролин и гидроксипролин (они не образуют достаточного количества водородных связей). Три α-цепи коллагена ориентированы параллельно и перевиты между собой, образуя субъединицу – тропоколлаген (микрофибриллу), представляющий собой плотный жгут:  Между полипептидными цепями микрофибрилл (тропоколлагена) образуются водородные, гидрофобные и ионные связи. Из тропоколлагена формируется четвертичная структура. Молекулы тропоколлагена сшиваются между собой поперечными ковалентными связями за счет аминогрупп радикалов лизина в более толстые коллагеновые фибриллы, а из них затем образуются волокна и пучки волокон: 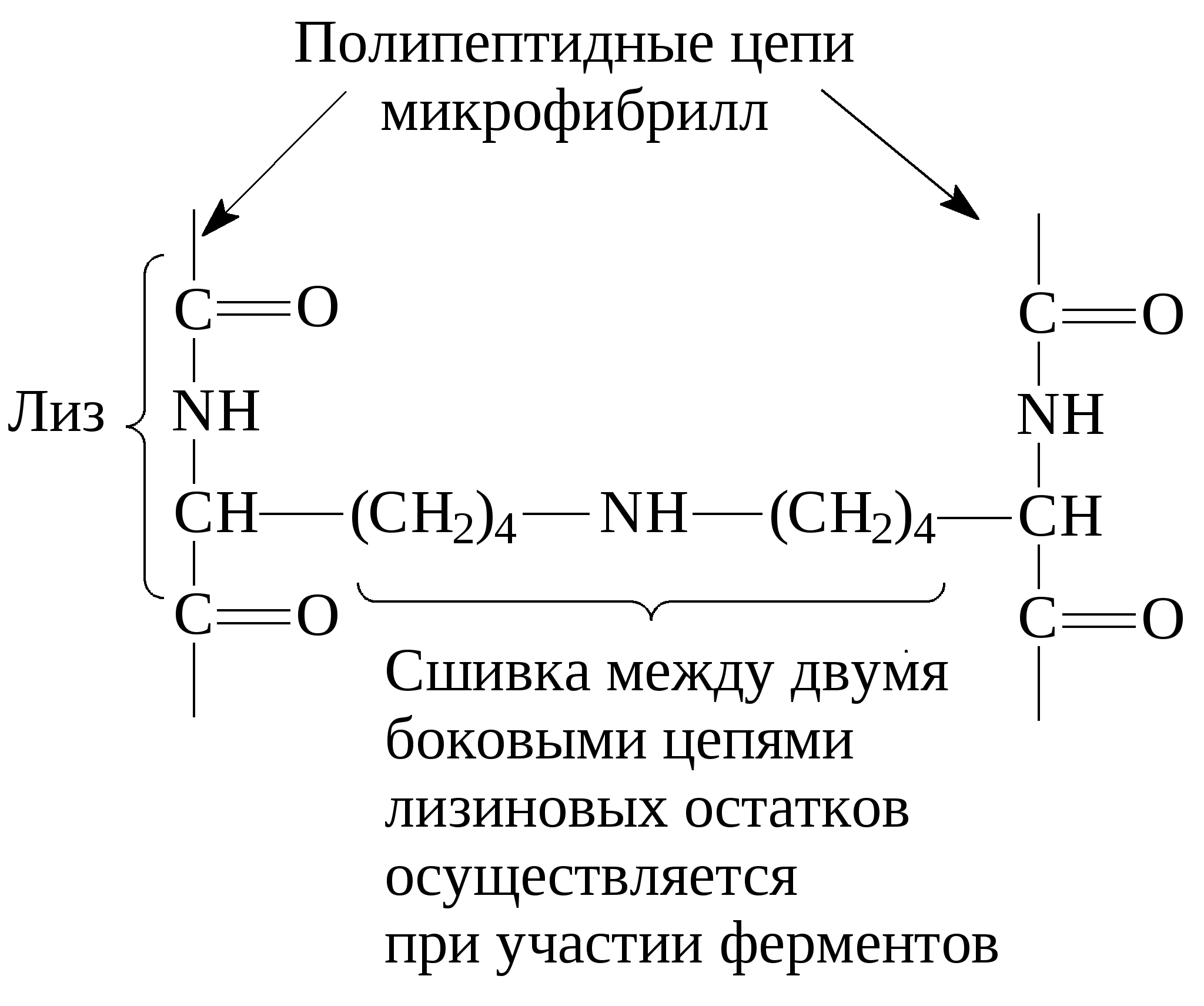 Тропоколлаген Тропоколлаген В ходе образования коллагена (созревания) многие остатки пролина и лизина гидроксилируются при участии аскорбиновой кислоты, которая выступает в качестве кофактора: 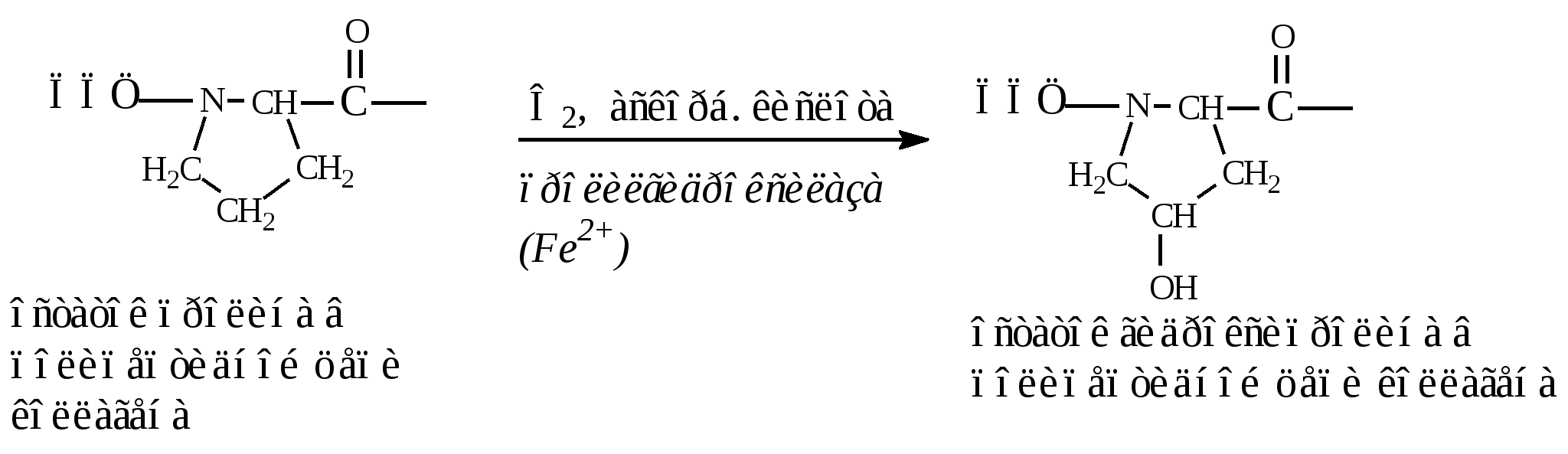 Витамин С необходим для поддержания в восстановленном состоянии ионов Fe2+ в активном центре фермента пролилгидроксилазы и для активации (восстановления) кислорода: При авитаминозе С нарушается формирование прочной структуры коллагена, что приводит к разрыхлению соединительной ткани. Помимо ковалентных свзей три спирально навитые друг на друга молекулы тропоколлагена в структуре коллагеновых фибрилл имеют дополнительную стабилизацию, которую формируют водородные связи при участии гидроксилированных остатков лизина и пролина. Например, сухожилия представляют собой пучки параллельно ориентированных коллагеновых фибрилл. В отличие от сухожилий, в коже коллагеновые фибриллы образуют подобие неупорядоченной двухмерной сетки. Пустоты в коллагеновых фибриллах могут служить местом первоначального отложения кристаллов гидроксиапатита – Са5(ОН)(РО4)3, играющего важную роль в минерализации костей. Известно несколько типов коллагенов, различающихся структурой полипептидных цепей, укладкой их во вторичную третичную и четвертичную структуры, степенью гликозилирования. Генетически обусловленные нарушения созревания коллагена после синтеза его полипептидных цепей на рибосомах приводит к тяжелым наследственным заболеваниям – коллагенозам. Все гликозаминогликаны (гетерополисахаридные цепи протеогликанов), содержащие уроновые кислоты и сульфатные группы при нейтральных значениях рН электростатически связываются с коллагеном. С каждым коллагеновым мономером связывается от 2 до 5 хондроитинсульфатных цепей протеогликанов. С эластином также специфически связываются хондроитинсульфаты и гепарансульфаты Эластин – содержится в больших количествах в межклеточном веществе соединительных тканей, испытывающих периодические растяжения и сокращения. Это гладкие мышцы крупных кровеносных сосудов, связок, легких. Например, в аорте эластин составляет 30–60% от массы вещества ткани, в выйной связке – 70–80%. Структура эластина менее изучена. Как и коллаген, эластин содержит много глицина (30%) и аланина (21%), но мало гидроксипролина и нет гидроксилизина. В его состав входят также лейцин (10%), лизин (10%), валин (13%) и другие гидрофобные аминокислоты. Эластин основной компонент эластических волокон соединительной ткани, резиноподобный полимер. Он содержит обычные α-спирали, но благодаря некоторым особенностям строения молекулы эластина обладают упругостью большей, чем резина. Эластин образует поперечно-сшитую сеть, которая своими необычными механическими свойствами обязана уникальному способу ковалентного связывания боковых цепей лизина: четыре сближенных лизиновых остатка ферментативно формируют так называемую десмозиновую структуру, объединяющую в один узел четыре участка полипептидных цепей:  Организованные таким образом полипептидные цепи тропоэластина могут соединяться в системы, способные обратимо растягиваться во всех направлениях. Эластин не растворяется в холодной и горячей воде, в солевых растворах, разведенных кислотах и щелочах. Даже крепкая серная кислота оказывает на него слабое действие. Из эластина не образуется желатин. Изоэлектрическая точка смещена в слабокислую сторону, поскольку незначительно преобладают кислые аминокислоты. В водной среде полипептидные цепи эластина принимают форму глобулы, которая окружена гидратной оболочкой. Гидрофобные радикалы аминокислот, образующие соответсвующие связи, спрятаны внутри белковой молекулы. Эластин не переваривается трипсином и химотрипсином, очень медленно гидролизуется пепсином в кислой среде при рН 2. В тонком кишечнике эластин расщепляется эластазой поджелудочной железы до аминокислот, пептидов и поперечно-сшитых пептидов. В пищевой промышленности для интенсификации процессов обработки и созревания мяса (коллагена, эластина, миозина) применяют ферментные препараты растительного (папаин, фицин, бромелаин) и микробного происхождения. 3.2. СЛОЖНЫЕ БЕЛКИ Структурная организация сложных белков представлена белковой частью (полипептидная цепь) и небелковым компонентом – простетической группой. В основе классификации сложных белков лежит характер простетчсекой группы. Гликопротеины и протеогликаны представляют собой углевод-белковые комплексы, простетическая группа которых включает олиго- и гетерополисахариды разнообразного строения. Их углеводная часть построена из моносахаридов (глюкозы, галактозы, маннозы и др.) и производных моносхаридов – гексозаминов (глюкозамин, галктозамин, маннозамин и др.), ацетилированных и сульфатированных гексозаминов, сиаловой (N-ацетилнейраминовой) и уроновой кислот, фукозы и др. Простетическая группа углевод-белковых комплексов может быть линейного и разветвленного строения, кислого или нейтрального характера. Этим белкам принадлежат многообразные функции в организме: рецепторная, защитная (иммунные белки), транспортная, они участвуют в структурной организации клеток и тканей. Липопротеины представляют собой липид-белковые комплексы, простетическая группа которых представлена простыми и сложными липидами (триацилглицеролами, холестерином, фосфо- и сфинголипидами и др.). Свободные липопротеины обладают гидрофильными свойствами ивыполняют транспортную функцию – транспортируют липиды в крови, молоке и других биологических жидкостях. Структурные липопротеины являются жирорастворимыми и входят в состав биомембран клеток и органелл клеток тканей животного происхождения. Нуклеопротеины сложные белки, простетическая группа которых представлена нуклеиновыми кислотами – дезоксирибонуклеиновая (ДНК) и рибонуклеиновая (РНК): ДНК-протеины и РНК-протеины. Белковый компонент представлен гистонами, протаминами или другими поликатионными белками. Они выполняют чрезвычайно важную роль в организме. ДНК-протеины хранят и передают генетическую информацию о структуре белка и всех видов РНК, а также служат матрицей для биосинтеза ДНК (при делении клетки) и всех видов РНК в процессе жизнедеятельности. РНК-протеины участвуют в синтезе белка в клетке. Хромопротеины – это сложные белки, простетическая группа которых имеет окраску. Большинство из них содержат металл. Наиболее распространенными в пищевых продуктах являются железосодержащие гемопротеины и магнийсодержащие хлорофиллпротеины. Витаминпротеины представляют собой сложные белки, простетическая группа которых представлена витаминами и витаминными кофакторами. Они выполняют каталитическую функцию, осуществляют обмен белков, углеводов, липидов, нуклеиновых кислот и энергетический обмен, участвуют в технологических процессах при производстве продуктов питания, при созревании мяса в послеубойный период и рыбы в посоле, сыроделии и др. процессах. Фосфопротеины представляют собой фосфорилированные формы белковых молекул. Важными представителями фосфопротеинов являются подфракции αS1-, αS2-, β- и χ-казеина молока. Остатки фосфорных кислот легко связывают ионы кальция Са2+ в молоке и, таким образом, казеин служит источником не только фосфора, но и кальция для организма человека. Многие белки в клетках и тканях находятся в фосфорилированной форме, например, ферменты. |