бх итоговая. БХ_итоговая_ласт. 1. Нервная и гуморальная регуляция как единая система регуляции обмена веществ. Гормоны первичные посредники в передаче информации
 Скачать 1.44 Mb. Скачать 1.44 Mb.
|

2. Регуляция синтеза и секреции гормонов по принципу обратной связи.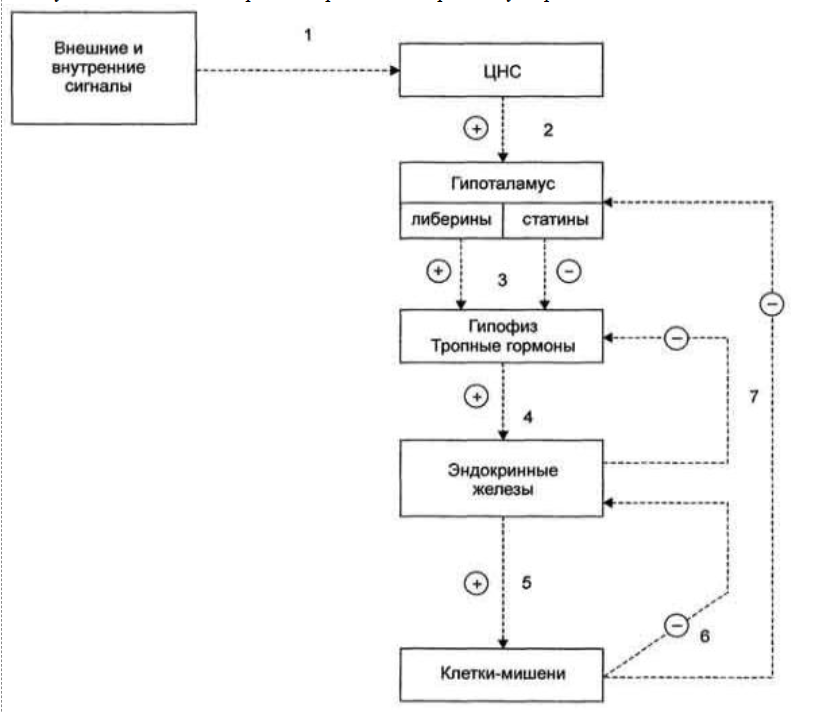 Под обратной связью подразумевается система, в которой конечный продукт деятельности этой системы (например, гормон, нейротрансмиттер и другие вещества) модифицирует или видоизменяет функцию компонентов, составляющих систему, направленную на изменение количества конечного продукта (гормона) или активности системы. Жизнедеятельность всего организма является следствием функционирования многочисленных саморегулируемых систем (выделительная, сердечно-сосудистая, пищеварительная, дыхательная и др.), находящихся в свою очередь под контролем нейроэндокринно-иммунной системы. Все перечисленное представляет, таким образом, комплекс различных саморегулируемых систем, находящихся в определенной степени зависимости и “подчиненности”. Конечный результат или активность системы может модифицироваться двумя путями, а именно путем стимуляции для увеличения количества конечного продукта (гормона) или повышения активности эффекта, или путем угнетения (ингибирования) системы с целью уменьшения количества конечного продукта или активности. Первый путь модифицирования называется положительной, а второй – отрицательной обратной связью. Примером положительной обратной связи является повышение уровня гормона в крови, стимулирующее высвобождение другого гормона (повышение уровня эстрадиола в крови вызывает высвобождение ЛГ в гипофизе), а отрицательной обратной связи, когда повышенный уровень одного гормона угнетает секрецию и высвобождение другого (повышение концентрации тироидных гормонов в крови снижает секрецию ТТГ в гипофизе). 3. Клетки-мишени и клеточные рецепторы гормонов. Строение рецепторов цитоплазматической мембраны (связанных с G-белками, с собственной ферментативной активностью). Рецепторы, локализованные в цитоплазме. Рецепторы сопряженные с ионными каналами. Регуляция работы рецепторного аппарата. Фактически все клетки животного организма являются мишенями для тех или иных гормонов. Истинная клетка-мишень — эта такая клетка, в которой при гормональном воздействии стимулируется специфическая биохимическая реакция клеточного метаболизма. Реализация эффекта зависит от концентрации гормона, взаимодействующего с клеткой, которая, в свою очередь, определяется скоростью биосинтеза гормона, созревания и условиями ассоциации-диссоциации с бслком-псрсносчиком в плазме крови. Конечный биохимический эффект зависит также от синергизма или антагонизма гормональных воздействий на клетки-мишени. Так, адреналин — гормон мозгового слоя надпочечников и глюкагон — гормон поджелудочной железы обладают сходным биохимическим действием: активацией распада гликогена в печени. Примером антагонистического действия могут служить эстрогены и прогестерон — женские половые гормоны, причем эстрогены усиливают сокращение матки, а прогестерон тормозит ее. Рецепторы, ассоциированные с G-белками – один из наиболее изученных классов мембранных рецепторов. Белки этой группы состоят из внеклеточного домена, семидоменной трансмембранной части и внутриклеточного домена. С такими рецепторами связан ассоциированный с мембраной адапторный белок с ГТФазной9 активностью (G-белок)10 (рис. 18). При связывании лиганда рецептор претерпевает конформационные изменения, передающиеся на G-белок. Тот, в свою очередь, воздействует на эффекторные молекулы, изменяя концентрации вторичных мессенджеров в цитоплазме. G-белки – гетеротримерные комплексы, состоящие из (рис. 18):  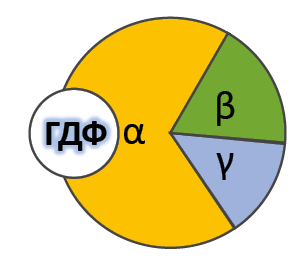 Ферментативной (ГТФазной) активностью обладает только α-субъединица G-белка. В неактивной форме она связана с ГДФ, но при взаимодействии с комплексом гормон-рецептор ГДФ заменяется на ГТФ, снижая сродство G-белка к рецептору. Из-за этого белок отсоединяется от рецептора и диссоциирует на α- и βγ-субъединицы11. Далее до гидролиза ГТФ α-субъединица успевает осуществить свою специфическую функцию, а после – вновь объединяется с βγ-субъединицами с образованием исходного G-белка (рис. 19). 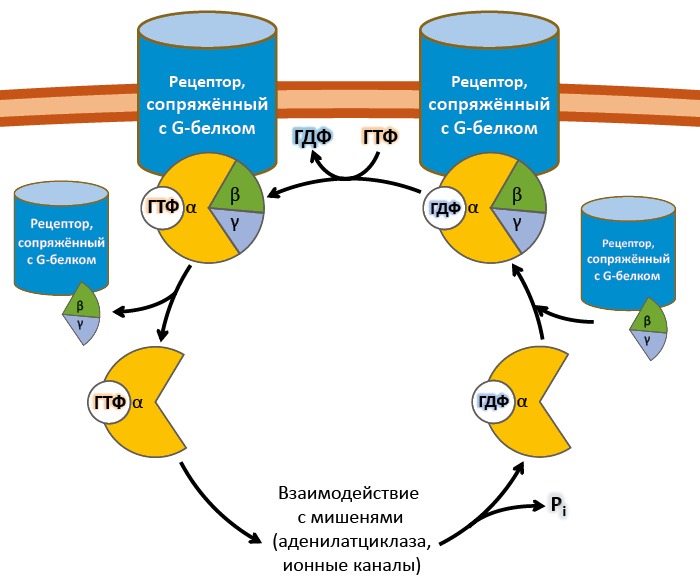 Рис. 19. Цикл активностиG-белков Несмотря на консервативность структуры и небольшую молекулярную массу, βγ-субъединицы не менее важны. В неактивном G-белке они связываются с α-субъединицей, удерживая её в неактивном (связанном с ГДФ) состоянии, и регулируют сродство рецептора к лиганду. А после диссоциации – активируют фосфолипазу А2, регулируют активность аденилатциклазы, ионных каналов и фосфорилирование рецепторов, обеспечивая «cross-talking» рецепторных систем. Так как β- и γ-субъединицы достаточно консервативны, классификация G-белков происходит на основании их α-субъединиц, определяющих специфическую функцию белкового комплекса. Выделяют следующие G-белки: Gs1 – активируют аденилатциклазу, L-Ca2+– каналы, ингибируют Na+-каналы; Gi1-3 – ингибируют аденилатциклазу, активируют K+– каналы и фосфолипазу C; Gq, G11, G14 – активируют фосфолипазу Сβ и Сδ; Gt и Gt1 – активируют фосфодиэстеразу; Gh – взаимодействуют с α-адренорецепторами; G12 – активирует систему малой ГТФазы Rho – протеинкиназ; Gx – предположительно активирует фосфолипазу С; G0 – предположительно активируют K+-каналы и фосфолипазу C, ингибируют Ca2+ – ПЗК12; p100 – предположительно участвет в эндоцитозе рецепторов. Для прикладной физиологии имеют значение три хорошо изученные группы G-белков, регулирующих основные сигнальные системы: Gs – активируют аденилатциклазную систему; Gi – ингибируют аденилатциклазную систему; Gq – активируют фосфолипазу С и фосфоинозитидную систему. G-белки не способны свободно передвигаться в цитоплазме. Они заякорены на внутренней поверхности цитоплазматической мембраны двумя углеводородными якорями: миристоевой кислотой (связывает α-субъединицу) и близкой холестерину геранил-гераниловой цепью (связывает βγ-субъединицы). Эти «якоря» одновременно обеспечивают и удержание белка на мембране, и его латеральную подвижность в плоскости мембраны. Несмотря на многообразие G-белков, их работа завязана на ограниченном наборе эффекторных систем клетки, регулируя которые, они осуществляют свою функцию. Сигнализация преимущественно осуществляется через аденилатцикланую и фосфоинозитидную системы. Виды мембраносвязанных рецепторов 1. Рецепторы, обладающие ферментативной активностью – при взаимодействии лиганда с рецептором активируется внутриклеточная часть (домен) рецептора, имеющий каталитическую (тирозинкиназную или тирозинфосфатазную или гуанилатциклазную) активность. По этому механизму действуют СТГ, инсулин, пролактин, интерлейкины, ростовые факторы, интерфероны α, β, γ. 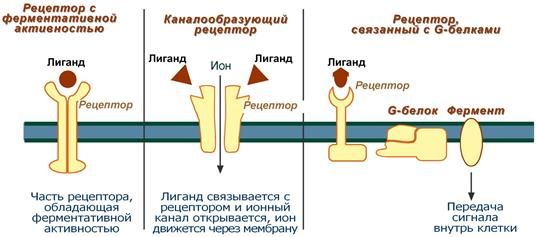 2. Каналообразующие рецепторы – присоединение лиганда к рецептору вызывает открытие ионного канала на мембране. Таким образом действуют нейромедиаторы – ацетилхолин (Na+-каналы), глицин и ГАМК (Cl¯-каналы), серотонин (Na+- и K+-каналы), глутамат (каналы для Na+, Ca+ и K+), вторичный мессенджер инозитолтрифосфат (Ca+-каналы). 3. Рецепторы, связанные с G-белками – передача сигнала от гормона происходит при посредстве особого G-белка. G-белок влияет на ферменты, образующие вторичные мессенджеры (посредники), которые уже передают сигнал на внутриклеточные белки. К третьему виду относятся аденилатциклазный и кальций-фосфолипидный механизмы. По данному механизму действуют большинство белковых и пептидных гормонов, некоторые медиаторы (серотонин, гистамин). 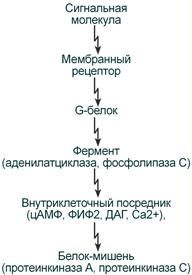 Схема механизма, связанного с G-белками  |