1 Определение жизнь с позиций системного подхода. Фундаментальные свойства живого
 Скачать 1.01 Mb. Скачать 1.01 Mb.
|

Этапы экспрессии гена эукариот: претранскрипционный, транскрипция, процессинг-сплайсинг, транспорт иРнк через ядерную мембрану, трансляция, посттрансляционныйДНК - про-иРНК - иРНК - белок Этапы экспрессии генов Претранскрипционный Транскрипция Процессинг и сплайсинг в ядре Транспорт иРНК через ядерную мембрану Трансляция Посттрансляционный этап В цитоплазме формирования функционально активного белка Претранскрипционный этап - активация генов. Активаторы: рН, ионы, БАВ, метаболиты и др. Регуляторы активности генов в претранскрипционный период: внутриклеточные (pH ±, Ионы ±,Белки (гистоновые, негистоновые) ±,Метаболиты ±, Медиаторы ±, БАВ ±, «Прыгающие гены» (их локализация)). Внеклеточные (Медиаторы ±, Гормоны ±,Др. раздражители). Все вызывают изменения МП плазм. мембр. ядерная МБР хромосомы. Функции регуляторов генной активности (в зоне действия): освобождают ДНК от белков, деконденсация ДНП, Ослабляют водородные связи, активируют РНК-полимеразу, блокируют белок-репрессор – освобождают оператор от блока (у прокариот), активируют регуляторные зоны. Транскрипция – синтез РНК на матрице ДНК, начинается с присоединения РНК-полимеразы к «своему» промотору (ТАТА-блоку); траскрипция происходит с одной полипептидной цепи ДНК (кодогенной), на которой находится промотор. Направление транскрипции 3’→5’. Нуклеотидная цепь иРНК растет в направлении 5’→3’.Транскрибируются все экзоны и интроны (включая зону копирования, кодон-инициатор, зоны терминации и полиаденилирования) структурного гена. В итоге образуется про-иРНК, содержащая кодирующие и некодирующие нуклеотидные последовательности. Фермент – РНК-полимераза. Процессинг про-иРНК: Кэпирование – образование КЭП на 5’-конце про-иРНК(служит для присоединения к рибосоме). Полиаденилирование на 3’-конце (присоединение поли-А или поли-У) - придает устойчивость иРНК. Процессинг – сплайсинг – «вырезание» интронов и «сшивание» (ферментами-рибозимами) экзонов. Транспорт иРНК через ядерную мембрану - транспорт зрелой иРНК через ядерную мембрану. Трансляция – синтез белков рибосомами на матрице иРНК. Зрелая иРНК 5’-концом (КЭП) подходит к полисомам и протягивается через них, транслируются кодон-инициатор и все нуклеотиды кодирующей зоны, кодоны-терминаторы не транслируются. Посттрансляционный этап - образование функционально активного белка: у эукариот «отрезается» метионин или триптофан, формируется вторичная, третичная, а для многих белков и четвертичная структура, присоединяются др. группировки и т.д. 14 - Уровни регуляции экспрессии генов. Механизмы и их характеристика, работающих на претранскрипционном и транскрипционном уровнях. Контроль экспрессии генов осуществляется на следующих уровнях: • на уровне транскрипции (контролируется время и характер транскрипции гена) • на уровне процессинга первичного транскрипта • при отборе зрелых мРНК для их транспорта в цитоплазму • на уровне трансляции - отбор в цитоплазме мРНК для трансляции на рибосомах • на уровне активности белка - селективная активация, инактивация или компартментация молекул белка после их синтеза. Уровни регуляции экспрессии генов 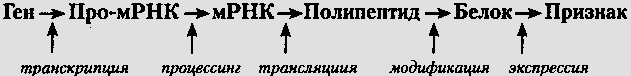 Претранскрипционный этап - активация генов. Активаторы: рН, ионы, БАВ, метаболиты и др. Регуляторы активности генов в претранскрипционный период: внутриклеточные (pH ±, Ионы ±,Белки (гистоновые, негистоновые) ±,Метаболиты ±, Медиаторы ±, БАВ ±, «Прыгающие гены» (их локализация)). Внеклеточные (Медиаторы ±, Гормоны ±,Др. раздражители). Все вызывают изменения МП плазм. мембр. ядерная МБР хромосомы. Функции регуляторов генной активности (в зоне действия): освобождают ДНК от белков, деконденсация ДНП, Ослабляют водородные связи, активируют РНК-полимеразу, блокируют белок-репрессор – освобождают оператор от блока (у прокариот), активируют регуляторные зоны. Транскрипция – синтез РНК на матрице ДНК, начинается с присоединения РНК-полимеразы к «своему» промотору (ТАТА-блоку); траскрипция происходит с одной полипептидной цепи ДНК (кодогенной), на которой находится промотор. Направление транскрипции 3’→5’. Нуклеотидная цепь иРНК растет в направлении 5’→3’.Транскрибируются все экзоны и интроны (включая зону копирования, кодон-инициатор, зоны терминации и полиаденилирования) структурного гена. В итоге образуется про-иРНК, содержащая кодирующие и некодирующие нуклеотидные последовательности. Фермент – РНК-полимераза. 16 - Мутации. Определение, классификация, механизмы возникновения МУТАЦИЯ — всеобщее свойство живых организмов, лежащее в основе эволюции и селекции всех форм жизни и заключающееся во внезапно возникающем изменении генетической информации. Для медицины изучение природы Мутаций чрезвычайно важно с точки зрения профилактики и лечения наследственных болезней 1. Различают спонтанные и индуцированные мутации. Спонтанные мутации возникают под действием естественных мутагенных факторов среды без вмешательства человека. Они увеличивают разнообразие живых организмов и создают материал для естественного отбора. Индуцированные мутации возникают при направленном воздействии на организм мутагенных факторов. Применение мутагенных воздействий позволяет увеличить количество мутаций в сотни раз. Так, использование селекционерами химических мутагенов позволило получить полиплоидные формы растений, которые отличаются устойчивостью к неблагоприятным условиям и большей продуктивностью. 2. Мутации могут возникать в соматических или половых клетках. Соматические мутации возникают в любых клетках, кроме гамет. Они затрагивают часть организма (например, разная окраска лепестков в одном цветке, разный цвет глаз у человека и животных). Такие мутации не наследуются при половом размножении, но передаются при вегетативном. Широко используются в селекции растений для выведения новых сортов. Генеративные мутации возникают в первичных половых клетках или в гаметах, передаются по наследству при половом размножении (например, гемофилия, синдром Дауна у человека). 3. По влиянию на жизнеспособность особей выделяют следующие виды мутаций: летальные (приводят к гибели мутантов); полулетальные (снижают жизнеспособность организма, вызывают наследственные заболевания, сокращают продолжительность жизни); нейтральные (изменяют признак, но не оказывают влияния на жизнеспособность организма); полезные (повышают жизнеспособность организма). 4. Мутации бывают доминантными и рецессивными. Доминантные мутации проявляются сразу и подвергаются действию естественного отбора (полезные сохраняются, вредные убираются). Большинство мутаций рецессивно, и проявиться они могут только в гомозиготном состоянии. Вероятность такого события мала, поэтому рецессивные мутации долгое время накапливаются в популяции в скрытом виде. 17 - Ген как единица изменчивости. Генные мутации и их классификация. Причины и механизмы возникновения генных мутаций. Ген-это единица наследственности и изменчивости. Каждый ген распологается в определенном участке хромосомы-локусе. Два гена находящиеся в одном локусе называются алельными. Изменчивость-свойство живых организмов приобретать в процессе онтогенеза отличия от родительских форм. Различают фенотипическую (не наследственную) и генотипическую(наследственную) изменчивость. Фенотипическая изменчивость не связанна с изменением генотипа и не сохраняется в следующем поколении, если нет условий, в которых она возникла(изменяется лишь функциональная активность генов). Фенотипическая изменчивость бывает возрастная и модификационная. Возрастная, или онтогенетическая выражается в изменении всего комплекса морфофизиологических и биохимических признаков организма на протяжении его индивидуальной жизни. Модификационная - возникает у генетически однородных особей одинакового возраста, которые находятся в разных условиях внешней среды, и проявляется в организме в качественных и количественных отклонениях. Диапазон модификационной изменчивости, в рамках которой зависимо от условий внеш.среды один и тот же генотип способен давать разные фенотипы, называют нормой реакции. Комбинативная изменчивость-связанна с новыми комбинациями генов в генотипе. Возникает такая изменчивость в результате процессов: 1.Независимого расхождения хромосом в мейозе( особый вид деления клеток, в результате которого образуются гаметы - половые клетки) 2.Случайного сочетания генов при оплодотворении 3.Рекомбинации генов благодаря перекрещиванию хромосом. Мутационная изменчивость- скачкообразное и устойчивое изменение генетического материала,передающегося по наследству. Возникает в следствии резких изменений признаков-мутаций и обусловливает реорганизацию структур воспроизведения, перестройку генетического аппарата. Этим мутации резко отличаются от модификации,которые не зацапают генотип особей. Мутации возникают внезапно, скачкообразно.образуя иногда формы организма , резко отменные от исходной формы. Генные мутации – изменение строения одного гена. Это изменение в последовательности нуклеотидов: выпадение, вставка, замена и т.п. Например, замена а на т. Причины – нарушения при удвоении (репликации) ДНК Мутация может заключаться: 1) в замене основания в кодоне, это так называемая миссенсмутация (от англ, mis - ложный, неправильный + лат. sensus - смысл) - замена нуклеотида в кодирующей части гена, приводящая к замене аминокислоты в полипептиде; 2) в таком изменении кодонов, которое приведет к остановке считывания информации, это так называемая нонсенсмутация (от лат. non - нет + sensus - смысл) — замена нуклеотида в кодирующей части гена, приводит к образованию кодона-терминатора (стоп-кодона) и прекращению трансляции; 3) нарушении считывания информации, сдвиге рамки считывания, называемом фреймшифтом (от англ. frame - рамка + shift: - сдвиг, перемещение), когда молекулярные изменения ДНК приводят к изменению триплетов в процессе трансляции полипептидной цепи. Известны и другие типы генных мутаций. По типу молекулярных изменений выделяют: делении (от лат. deletio - уничтожение), когда происходит утрата сегмента ДНК размером от одного нуклеотида до гена; дупликации (от лат. duplicatio - удвоение), т.е. удвоение или повторное дублирование сегмента ДНК от одного нуклеотида до целых генов; инверсии (от лат. inversio - перевертывание), т.е. поворот на 180° сегмента ДНК размерами от двух нукпеотидов до фрагмента, включающего несколько генов; инсерции (от лат. insertio - прикрепление), т.е. вставка фрагментов ДНК размером от одного нуклеотида до целого гена. -Генные мутации могут происходить в результате: 1) замены одного или нескольких нуклеотидов на другие; 2) вставки нуклеотидов; 3) потери нуклеотидов; 4) удвоения нуклеотидов; 5) изменения порядка чередования нуклеотидов. Эти мутации приводят к изменению аминокислотного состава полипептидной цепи и, следовательно, к изменению функциональной активности белковой молекулы. Благодаря генным мутациям возникают множественные аллели одного и того же гена. 18 - Хромосомы. Определение. Структурная организация хроматина. Морфология хромосом. Нуклеосомная модель строения хромосом Хромосомы— нуклеопротеидные структуры клетки, в которых сосредоточена большая часть наследственной информации и которые предназначены для её хранения, реализации и передачи. Хромосомы чётко различимы в световом микроскопе только в период митоза или мейоза. Набор всех хромосом клетки, называемый кариотипом. Структурная организация хроматина. Хроматин в зависимости от периода и фазы клеточного цикла меняет свою организацию. В интерфазе он выявляется в виде глыбок, рассеянных в нуклеоплазме ядра(в ядерном соке). При переходе клетки к митозу, особенно в метафазе, хроматин приобретает вид интенсивно окрашенных телец — хромосом.Интерфазную и метафазную формы существования хроматина рассматривают как два различных состояния одного и того же вещества, связанных в митотическом цикле взаимопереходами,и в основе обеих эти форм лежит одна и та же элементарная нитчатая структура. Т.е хроматин представляет собой спирализованную нить. Уровни компактизации (спирализации) хроматина. Первый уровень спирализации–образование двойной спиралиДНК, построенной по принципу комплементарности. (открыт впервые Уотсоном и Криком) Второй уровень (упаковка молекулы ДНК в нуклеосомную нить с помощью гистоновых и негистоновых белков).Нуклеосомная нить. Этот уровень организации хроматина обеспечивается четырьмя видами нуклеосомных гистонов: Н2А, Н2В, НЗ, Н4. Они образуют напоминающие по форме шайбу белковые тела — коры, состоящие из восьми молекул (по две молекулы каждого вида гистонов) Молекула ДНК соединяется с белковыми корами, спирально накручиваясь на них. При этом в контакте с каждым кором оказывается участок ДНК, состоящий из 146 пар нуклеотидов (п.н.). Свободные от контакта с белковыми телами участки ДНК называют связующими или линкерными. Они включают от 15 до 100 п.н. (в среднем 60 п.н.) в зависимости от типа клетки.Отрезок молекулы ДНК длиной около 200 п. н. вместе с белковым кором составляет нуклеосому. Благодаря такой организации в основе структуры хроматина лежит нить, представляющая собой цепочку повторяющихся единиц — нуклеосом . Вдоль нуклеосомной нити, напоминающей цепочку бус, имеются области ДНК, свободные от белковых тел. Эти области, расположенные с интервалами в несколько тысяч пар нуклеотидов, играют важную роль в дальнейшей упаковке хроматина, так как содержат нуклеотидные последовательности, специфически узнаваемые различными негистоновыми белками.В результате нуклеосомной организации хроматина двойная спираль ДНК диаметром 2 нм приобретает диаметр 10—11 нм Третий уровень.Хроматиновая фибрилла(нуклеомерный уровень).Дальнейшая компактизация нуклеосомной нити обеспечивается гистоном Н1, который сближает белковые коры. В результате образуется более компактная структура, построенная, возможно, по типу соленоида. Такая Хроматиновая фибрилла, называемая также элементарной, имеет диаметр 20—30 нм Четвертый уровень.Интерфазная хромонема(хромомерный). Скручивание самой нуклеосомной нити приводит к образованию элементарной хроматиновойфибриллы.Каждая хроматида состоит из одной фибриллы. Пятый уровень (метафазная хроматида) – при дальнейшей упаковке хроматиновые фибриллы образуют петельные домены, внутри которых встречаются более конденсированные участки. Таким образом, упакованная хроматиновая фибрилла образует хроматиду, а две хроматиды одну хромосому. Общая морфология хромосом лучше всего выявляется на стадии метафазы и ранней анафазы, когда хромосомы наиболее укорочены и находятся в экваториальной плоскости. В метафазе и анафазе хромосомы одной клетки различаются по форме Типы хромосом определяются главным образом положением первичной перетяжки хромосомы, где располагается центромера. Кроме первичной перетяжки, хромосома может иметь вторичную перетяжку, не имеющую отношения к прикреплению нитей веретена. Местонахождение этой перетяжки в хромосоме связано с формированием ядрышка. Этот участок хромосомы называется ядрышковым (нуклеолярным) организатором. Полагают, что он имеет сложную структуру и ответствен за синтез рибосомной РНК. Иногда на концах хромосом находятся небольшие тельца — спутники. Такие хромосомы называются спутничными. Каждая хромосома обязательно имеет центромеру, выполняющую функцию механического центра хромосомы. Именно к ней в метафазе прикрепляется нить веретена, разводящая хромосомы к полюсам. Местоположение центромеры в разных хромосомах может быть различным, но оно типично для каждой хромосомы. Если центромера располагается в хромосоме посередине, то в метафазе хромосома выглядит как равноплечая V-образная, или метацентрическая. Если центромера делит хромосому на два неравных участка, то образуются или слабо неравноплечая — субметацентрическая или резко неравноплечая — акроцентрическая хромосомы. Размещение центромеры у конца хромосомы делает ее в метафазе палочкообразной, или телоцентрической. Предполагают, что центромера никогда не бывает на самом конце хромосомы. Участок хромосомы, располагающийся ближе к центромере, называют проксимальным, а отдаленный — дистальным. Центромеры определяют ориентацию хромосом в митозе и их правильное расхождение к полюсам. Если тонким пучком ультрафиолетовых лучей облучить участок хромосомы с центромерой, то хромосома теряет эту ориентацию. В случае разлома хромосомы и потери центромеры бесцентромерный ее участок (ацентрический фрагмент) восстановить центромеру не может. В силу отсутствия центромеры такой ацентрический фрагмент не может распределяться нормально при клеточном делении и чаще всего утрачивается. Фрагмент сохранится лишь в случае, если он прикрепится к хромосоме, имеющую центромеру. Центромера содержит ДНК и также является самовоспроизводящейся структурной частью хромосомы. Известны короткие палочковидные хромосомы, которые по всей своей длине или в значительной части обладают функцией центромеры — они имеют так называемую диффузную центромеру. Такие полицентрические хромосомы имеют много нитей веретена, прикрепляющихся к хромосоме по всей ее длине. В этом случае даже разорвавшиеся хромосомы могут нормально расходиться в анафазе. Природа этого явления остается пока мало исследованной. На концах хромосом имеются сегменты, препятствующие склеиванию хромосом концами. Такие сегменты названы теломерами. |