1. Введение Жизнь на Земле представляет собой форму существования материи
 Скачать 28.03 Mb. Скачать 28.03 Mb.
|

9. Царство грибыMycota, Fungi Грибы – одно из самых больших царств организмов. Появились первые представители 450 – 500 млн. лет назад. Их группы произошли независимо, от разных жгутиковых. Некоторые предположительно произошли от водорослей или амебоидных предков. Существует около 120000 видов грибов, разнообразных по внешнему виду и распространению. По наличию в обмене веществ мочевины, хитина в клеточной стенке клеток, запасного продукта – гликогена грибы приближаются к животным. А по способу питания путем всасывания (абсорбтивное питание), а не заглатывания пищи, и по неограниченному росту они напоминают растения. Общая характеристика, особенности строения, питания, размножения: Тело гриба представляет собой мицелий (грибницу), который состоит из разветвленной сети тонких нитей – гиф. У грибов, условно называемых низшими, мицелий неклеточный (не имеет перегородок) и представляет собой одну разветвленную клетку; у остальных - гифы разделены перегородками (септами) на отдельные клетки, образуя многоклеточный мицелий. В септах многоклеточного мицелия имеются отверстия (поры), через которые осуществляется цитоплазматическая связь между клетками. Кроме того, гифы мицелия могут сильно переплетаться, образуя ложную ткань – плектенхиму. У некоторых грибов – паразитов мицелий отсутствует, а тело представляет собой плазмодий. Клетки имеют клеточную стенку и протопласт. Клеточная стенка у низших грибов состоит из пектиновых веществ, у высших – из углеводов близких к целлюлозе, со значительной примесью хитина (близок к хитину насекомых). В протопласте выделяется цитоплазма (у молодых клеток гомогенна), в которой хорошо различима плазмалемма и тонопласт, рибосомы, митохондрии, аппарат Гольджи, эндоплазматическая сеть, 1-2 или много мелких ядер. Пластид нет. Запасные вещества – гликоген, жиры, волютин (находятся в цитоплазме), гранулы белков (в вакуолях). Крахмала нет. Грибы – гетеротрофы (питаются готовыми органическими веществами). По образу жизни их делят на две группы. Первая – сапротрофы (получают органические вещества из органических останков), вторая – паразиты (используют живую субстанцию растений, животных, человека и др. в качестве источника органических веществ). К первой группе относится большинство грибов. Питание грибов происходит путем осмоса всей поверхностью мицелия, реже при помощи всасывающих ризоидов (плесневые грибы) или присосок – гаусторий у паразитов (ржавчинные, головневые грибы). Размножаются грибы вегетативным, бесполым и половым путем. Вегетативное размножение грибов происходит: 1) частями мицелия; 2) оидиями (артроспорами) - образуются в результате распадения гиф на отдельные клетки, дающие новый мицелий; 3) хламидоспорами (образуются как оидии) - имеют более твердую, часто темноокрашенную оболочку (головневые грибы) и предназначены для перенесения неблагоприятных условий; 4) почкованием - на мицелии образуются боковые выросты, которые отчленяются от него. Бесполое размножение у грибов происходит тремя способами: зооспорами, спорангиоспорами и конидиями. Зооспоры (преимущественно у водных грибов) имеют жгутики, подвижны и образуются в зооспорангиях. Спорангиоспоры – это неподвижные споры, которые развиваются в спорангиях. Конидии характерны для высших грибов (аскомикоты, базидиомикоты, дейтеромикоты) и образуются на особых гифах – конидиеносцах. При этом кончик конидиеносца отделяется перегородкой, округляется и отпадает в виде конидии. Под первой конидией до ее отделения таким же образом формируется следующая и т.д., в результате образуется целая цепочка. Половое размножение у грибов очень разнообразно и отмечено у всех таксонов, кроме дейтеромикотов. Причем половой процесс состоит из двух этапов: 1 - слияние цитоплазмы двух клеток (плазмогамия); 2 - слияние ядер (кариогамия), и эти этапы разделены во времени. Различают несколько типов полового процесса (рис. 9.1). 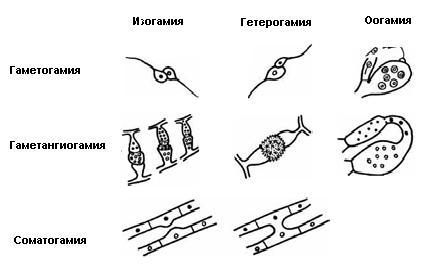 Рис. 9.1. Некоторые типы полового процесса у грибов. гаметогамия – слияниие гамет, образующихся в гаметангиях; гаметангиогамия - слияние содержимого двух многоядерных специализированных половых органов (мужского и женского гаметангиев), которые не дифференцированы на гаметы; в качестве гамет здесь можно рассматривать свободно располагающиеся в гаметангиях ядра; характерен для зигомикотов и аскомикотов; соматогамия - половых органов не образуется, а сливаются гаплоидные соматические клетки физиологически различных гиф; характерен для базидиомикотов; хологамия (у грибов с одноклеточным мицелием) - наблюдается слияние двух особей. Половой процесс у так называемых высших грибов завершается образованием зиготы, ее мейотическим делением и образования полового спороношения. В систематике грибов еще очень много спорного. До недавнего времени существовало деление грибов на низшие (хитридиомицеты, оомицеты и зигомицеты) и высшие (аскомицеты, базидиомицеты и дейтеромицеты). В настоящее время, к настоящим грибам относят четыре таксона, которые имеют общее происхождение – зигомикоты, аскомикоты, базидиомикоты, дейтеромикоты (несовершенные грибы) и лишайники, слоевища которых образованы мицелием грибов. Оомикоты и Хитридиомикоты отнесены к грибоподобным протоктистам. Перечисленные таксоны обычно считают классами (имеют окончания - мицета, -mycetes) или отделами (имеют окончание - микота, - mycota). Отдел Зигомикоты (Zygomycota) Содержит более 500 видов, ведущих наземный образ жизни, среди которых есть и сапротрофы, и паразиты. Представители этого отдела имеют хорошо развитый нечленистый мицелий (не разделен перегородками и представляют собой одну разветвленную клетку). Бесполое размножение - неподвижными спорами. Половой процесс состоит в слиянии двух, чаще внешне не различимых клеток на концах мицелия (зигогамия). У многих из них сливаться могут лишь клетки, имеющие разные половые знаки, условно обозначаемые + или -, хотя внешне и одинаковые. Это явление названо гетероталлизмом (раздельнополость). Открыто оно было у мукоровых грибов, а в настоящее время известно у грибов из многих систематических групп. Один из представителей данного отдела Мукор, или головчатая плесень (Mucormucedo) – сапротроф. Развивается на увлажненном хлебе, плодах, овощах, навозе в виде белой пушистой плесени. От мицелия приподнимаются вертикальные спорангиеносцы со спорангиями (вначале булавовидной, а затем шаровидной формы), внутри которых образуются споры. Совокупность спорангиеносцев образует обычно пушок серого или белого цвета с бурыми или черными головками на концах. Споры разносятся по воздуху и в благоприятных условиях образуют новый мицелий. Половой процесс наблюдается лишь при встрече и соприкосновении двух разнополых гиф. На концах таких гиф образуется по крупной многоядерной клетке, ограниченной перегородкой. В месте соприкосновения разнополых гиф в результате расслизнения стенок клеток происходит слияние цитоплазмы и ядер обеих клеток. Образовавшаяся зигота с многочисленными диплоидными ядрами покрывается толстой бурой шиповатой оболочкой. После длительного периода покоя ядра претерпевают мейоз, и зигота прорастает в зародышевый спорангий, внутри которого образуются споры, дающие разнокачественные гифы (рис 9. 2). 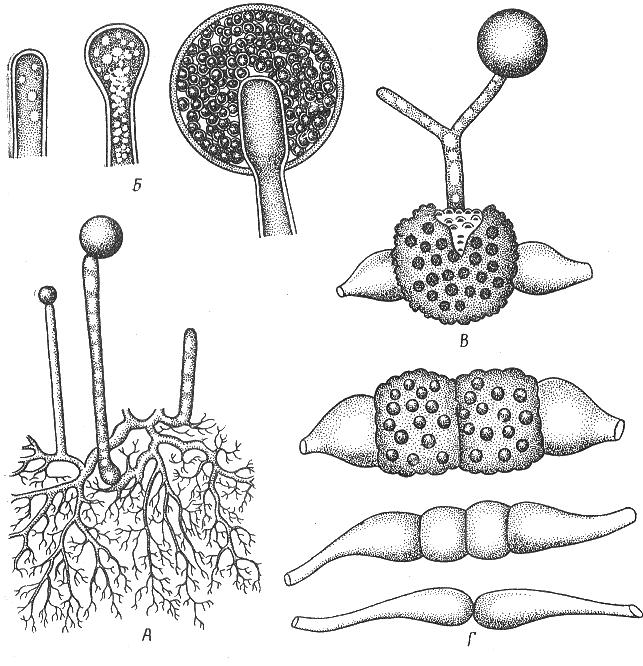 Рис. 9. 2. Мукор (Mucormucedo): А – мицелий со спорангиями; Б – образование спорангия; В – прорастание зиготы; Г – гаметангиогамия. Отдел Аскомикоты, или сумчатые грибы (Ascomycota) Включает более 30 тыс. видов. Среди них есть как сапротрофы, так и биотрофы (паразиты и симбионты); одноклеточные организмы (дрожжи) и виды с клеточным, хорошо развитым мицелием, образующим плодовые тела до 10- 20 см (сморчки, строчки, трюфели). Размножаются сумчатые грибы спорами, которые образуются в асках (сумках). Образованию спор предшествует половой процесс, протекающий в скрытой форме. На гифах мицелия образуются органы полового размножения (архикарпы – женские, антеридии – мужские), содержащие цитоплазму и несколько ядер. Архикарп состоит из нитевидной трихогины и расширенного основания – аскогона, антеридий – одноклеточное образование овальной формы. При соприкосновении этих органов трихогина врастает своим концом в полость антеридия и содержимое антеридия перемещается в аскогон, далее обе цитоплазмы сливаются (плазмогамия), а ядра располагаются попарно, образуя дикарионы. Оплодотворенный аскогон прорастает аскогенными гифами. Конечные клетки последних разрастаются, в них происходит слияние ядер дикариона (кариогамия) с образованием одного диплоидного ядра. Это ядро делится сначала путем мейоза, затем митоза и образуется 8 гаплоидных ядер будущих спор. В результате на конце аскогенных гиф развиваются сумки, содержащие аскоспоры (рис. 9. 3). 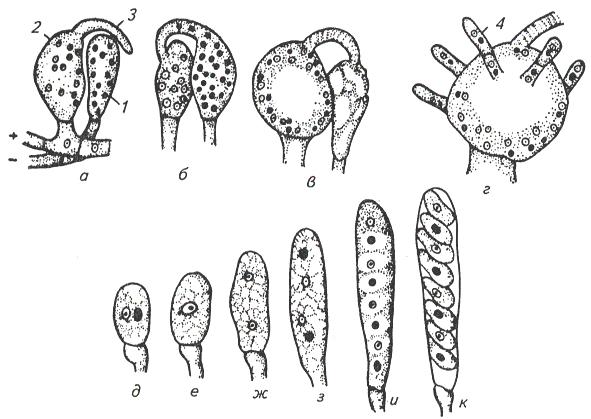 Рис. 9. 3. Половой процесс у сумчатого гриба пиронемы: а – антеридий (1) и архикарп (2) с трихогиной (3); б – переход содержимого антеридия в архикарп; в – образование в аскогоне дикарионов; г – образование дикарионных аскогенных гиф (4); д, е – образование зиготы на верхушке аскогенной гифы; ж, з, и, к – мейоз и развитие сумки с аскоспорами. Кроме полового процесса, у аскомикотов есть вегетативное размножение (частями мицелия, оидиями, почкованием) и бесполое – с помощью конидий, развивающихся на конидиеносцах. Отдел аскомикоты делится на классы: голосумчатые, плодосумчатые. Класс Голосумчатые грибы (Hemiascomycetes). Сумки развиваются непосредственно на мицелии, плодовых тел нет. Так виды рода Saccharomyces (дрожжевые грибы) представляют собой одиночные, одноядерные, сферические или овальные клетки, которые размножаются вегетативно почкованием. Последнему способствует достаточное количество сахара, температура – 25-30°С и аэрация. Только в результате интенсивного размножения возникают цепочки клеток (псевдомицелий), которые в последствии распадаются (рис. 9. 4). 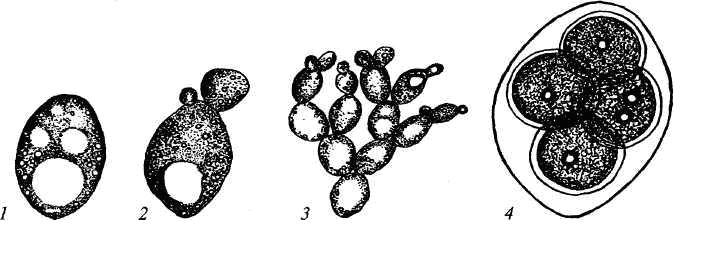 Рис. 9. 4. Хлебные дрожжи ( Saccharomyces cerevisiae): 1 – клетка дрожжей, 2 – начало почкования, 3 – почкование (образоваие псевдомицелия), 4 – сумка со спорами. При истощении питательной среды у дрожжей можно наблюдать половой процесс. Он состоит в слиянии (копуляции) двух гаплоидных клеток. Впоследствии из зиготы образуется сумка с 4-8 аскоспорами. Рассматриваемый род объединяет как природные виды, так и «культурные» (производственные), существование которых тесно связано со всей историей бродильной промышленности. Большое практическое значение имеют пивные, или хлебные дрожжи ( Saccharomyces cerevisiae), которые существуют только в виде культурных рас и винные дрожжи ( S. vini, S. ellipsoideus), которые живут на поверхности и в соке различных плодов. Колонии дрожжевых грибов выглядят следующим образом (рис. 9. 5). 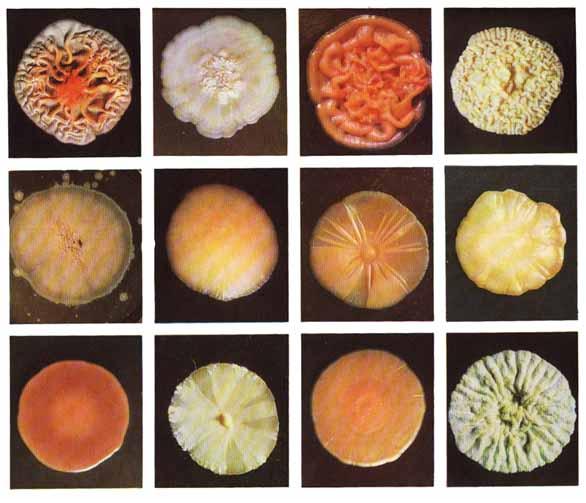 Рис. 9. 5. Колонии дрожжевых грибов . Другие представители данного класса (например виды рода Taphrina – тафрина), вызывают поражение стеблей, листьев, цветков, плодов растений, способствуют их разрастанию и деформации. Класс Плодосумчатые грибы (Carpoascomycetes). У представителей этого класса сумки развиваются в специальных вместилищах – плодовых телах, различных по форме и величине. Существует несколько типов плодовых тел: клейстотеций (замкнутое, шарообразное плодовое тело), перитеций (полузамкнутое, кувшиновидное плодовое тело с отверстием на вершине) и апотеций (открытое, чашевидное плодовое тело). Представитель данного класса - Спорынья (Claviceps) паразитирует на злаках (рожь), осоках. Имеет плодовое тело перитеций. В конце лета в колосьях ржи вместо зерновок образуются черно-фиолетовые рожки (склероции), которые состоят из плотного переплетения гиф. Цикл развития спорыньи Включает три стадии: склероциальную, сумчатую и конидиальную. Стадия I - образование склероция (покоящейся стадии гриба). При сильном поражении ржи на отдельных колосьях может быть до 3-4 склероциев. Далее при уборке хлеба склероции могут самопроизвольно опадать на землю (они хорошо переносят морозы и на следующий год после всходов ржи начинают прорастать), или при обмолоте попадать в товарное или семенное зерно. Стадия II- на прорастающем склероции появляются красные или темно-розовые булавовидные плодовые тела, состоящие из тонких ножек и шаровидных головок, усаженных многочисленными мелкими коническими выступами ("бородавочками"). Эта стадия – строма. Бородавочки на головке являются выходами перитециев - яйцевидных полостей, образующихся в периферической части головки. В перитециях вырастают многочисленные булавовидной формы сумки, в каждой из которых развивается по 8 нитевидных аскоспор. К моменту цветения ржи плодовые тела гриба полностью созревают; при этом из слизисто разбухающих перитециев выдавливаются сумки, которые лопаются; при этом из них выбрасываются аскоспоры и воздухом разносятся по цветущей ржи. Стадия III начинается с попадания аскоспор на перистые рыльца цветков ржи и их прорастания. Из сплетения гиф на завязи цветка образуется грибница, по мере развития которой начинается бесполое размножение гриба. Заключается оно в отшнуровывании с концов гиф многочисленных мелких эллиптических конидиоспор. Одновременно грибницей вырабатывается клейкая жидкость, содержащая сахаристые вещества, называемая "медвяной росой". Капли последней стекают по пораженному колосу, унося с собой конидиоспоры. Сладкая жидкость привлекает насекомых, которые, перелетая на другие колосья, разносят конидиоспоры, способствуя, тем самым, новому (повторному) заражению ржи. Конидиоспоры, попав на здоровые цветки ржи, также прорастают, образуя на завязи грибницу. Постепенно грибницы (образовавшиеся как из аскоспор, так и из конидиоспор), разрастаясь, разрушают завязь, и, в конечном счете, на месте и вместо зерна развивается белое продолговатое крупное грибное тело - молодой склероций. К моменту созревания ржи созревают и склероции; гифы уплотняются, наружный слой склероция при этом пигментируется, окрашиваясь в темно-фиолетовый цвет (рис. 9. 6 (а) - 9. 6 (б)). 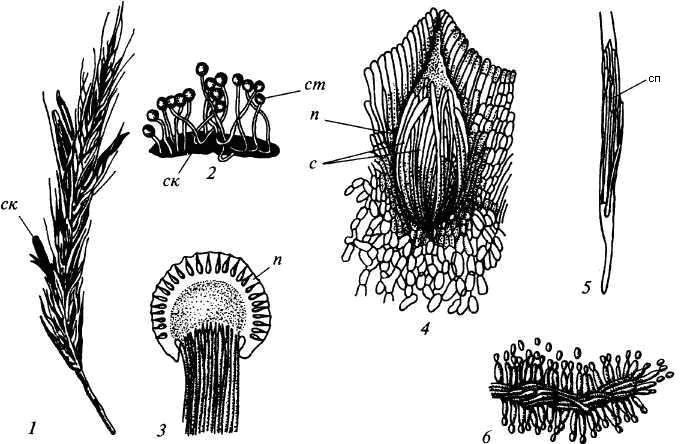 Рис. 9. 6 (а). Спорынья пурпурная (Claviceps purpurea): 1 – колос ржи со склероциями, 2 – склероций, проросший головчатыми стромами, 3 - разрез стромы с перитециями, 4 – отдельный перитеций в строме с сумками, 5 – сумка с аскоспорами, 6 – конидиальное спороношение, ск – склероции, ст – стромы, п – перитеций, с – сумки, сп – споры.  Рис. 9. 6 (б). Спорынья пурпурная (Claviceps purpurea): колос ржи со склероциями, склероций, проросший головчатыми стромами. Рожки (склероции) спорыньи эрготаминового (эрготоксинового) штамма ( Cornua Secalis cornuti stamm Ergotamini (Ergotoxini)) содержат алкалоиды индольного ряда, обладающие ядовитыми свойствами и оказывающие сложное влияние на организм человека. Незначительное количество размолотых склероцией в муке способно вызвать тяжелое заболевание – эрготизм, иногда приводящее к смерти. В современной медицине алкалоиды спорыньи широко применяются для лечения сердечно-сосудистых и нервных заболеваний (адреноблокирующая активность), а также в акушерской практике (вызывают сокращение матки). Плодовое тело - апотеций имеют пецица (Peziza), строчок (Gyromitra), сморчок (Morchella) (рис. 9. 7). 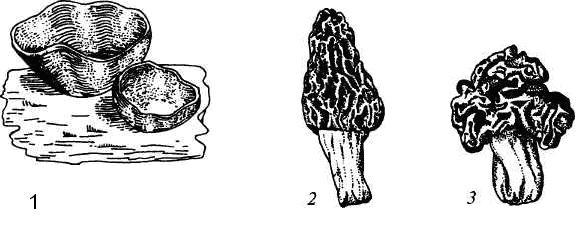 Рис. 9. 7 (а). Апотеции пецицевых : 1 – пецица, 2 - сморчок, 3 – строчок. 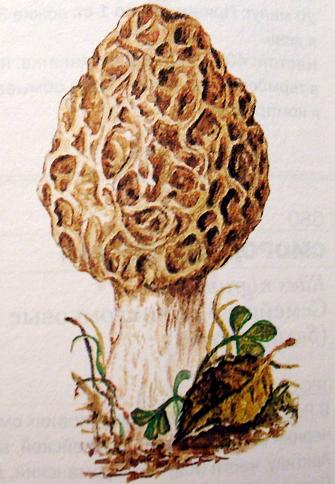 Рис. 9. 7 (б). Сморчок настоящий. Пецица встречается в лесах, на местах пожарищ, на почве огородов, коровьем навозе. Ее апотеции более или менее мясистые, и имеют окраску от желтой до красной и коричневой. Строчок и сморчок встречаются в лесу рано весной. Плодовое тело состоит из шляпки с морщинистой поверхностью, выстланной гимением (слой асков, разделенных парафизами (бесплодными гифами)) и ножки. Строчки содержат ядовитую гельвелловую кислоту (разрушается после длительного кипячения). Отдел Базидиомикоты, или базидиальные грибы (Basidiomycota) Насчитывает около 30 тыс. видов. Представители этого отдела имеют многоклеточный (септированный) мицелий. Среди базидиомикот многочисленные сапротрофы и опасные паразиты высших растений. Значительное число видов – микоризообразователи (микориза – симбиотическая ассоциация гриба с корнями растений). Конидиальное спороношение у базидиомикот встречается редко. Половой процесс осуществляется путем слияния двух вегетативных клеток гаплоидного (первичного) мицелия. Специальных органов полового размножения у базидиомикот нет. Органом полового спороношения является особая репродуктивная структура - базидия, на которой образуются базидиоспоры. Гифы, вырастающие из базидиоспор, гетероталличны (берут начало от спор противоположных половых знаков: «+» и «-»), и при их соприкосновении происходит половой процесс (соматогамия). При этом содержимое клетки одной гифы переходит в клетку другой, где происходит слияние цитоплазмы (плазмогамия). Ядра не сливаются, а образуют пары – дикарионы, которые впоследствии одновременно делятся, образуя дикарионный (вторичный) мицелий. На дикарионном мицелии образуются выросты – базидии – куда переходят дикарионы с цитоплазмой. В базидии завершается половой процесс: сливаются ядра дикариона (кариогамия), редукционно (мейозом) делится диплоидное ядро и возникает 4 гаплоидных ядра. В верхней части базидии образуются четыре трубчатых выроста с расширением на конце – стеригмы. В них переходят ядра с цитоплазмой и возникают четыре базидиоспоры: две со знаком «+» и две со знаком «-», впоследствии образующие гетероталличные гаплоидные мицелии (рис. 9. 8). 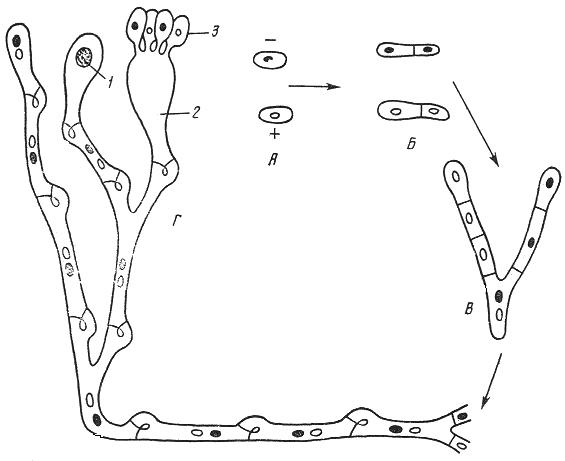 Рис. 9. 8. Половой процесс у базидиомикот (Basidiomycota): А – базидиоспоры; Б – гаплоидные гифы; В – соматогамия; Г – формирование базидии: 1 – зигота, 2 – базидия, 3 – базидиоспора. Различают три типа базидий: холобазидии (одноклеточные, булавовидные или цилиндрические), гетеробазидии состоят из двух частей (гипобазидии и эпибазидии), фрагмобазидии (разделены перегородками на 4 клетки, по бокам которых формируются базидиоспоры) (рис. 9. 9). 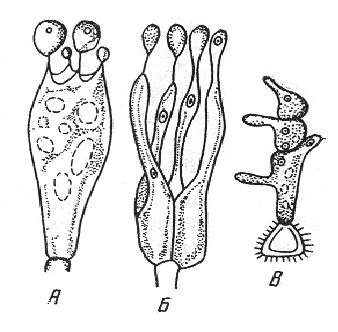 Рис. 9. 9. Типы базидий: А – холобазидия; Б – гетеробазидия; В – фрагмобазидия. Базидии с базидиоспорами могут возникать прямо на мицелии, либо на плодовых телах (или внутри них). Плодовые тела различны по форме и консистенции (рыхлые, паутинистые, деревянистые и т.д.). На их верхней или нижней стороне располагается спороносный слой – гимений. Поверхность плодового тела, несущая гимений, называется гименофором. По типу развития и строению базидии базидиомикоты подразделяются на три класса: холобазидиомицеты (Holobasidiomycetes), фрагмобазидиомицеты (Phragmobasidiomycetes), гетеробазидиомицеты (Heterobasidiomycetes). Класс Холобазидиомицеты (Holobasidiomycetes).Это, в основном, сапрофиты. Базидии одноклеточные и вместе с бесплодными гифами образуют гимениальный слой. Последний развивается на гименофоре (плотная основа плодового тела из сплетенных гиф), который может быть трубчатым и пластинчатым. Трубчатый гименофор имеют представители семейств трутовиковые и болетовые, пластинчатый – грибы из семейств пластинниковые, мухоморовые. К семейству трутовиковые относятся: домовой гриб (Serpulalacrymans) - разрушитель древесины; настоящий трутовик (Fomesfomentarius) - паразитирует на березе, буке, имеет многолетнее плодовое тело копытообразной формы; ложный трутовик (Phellinusigniarius) - паразитирует на многих лиственных деревьях (ольха, осина, береза, дуб), вызывает образование дупел; трутовик косотрубчатый (чага) (Inonotusobliquus) - повсеместно поражает березовые леса, образуя черные растрескивающиеся наросты на березе. Чагу заготавливают для медицинских целей, готовят настойки и экстракты, обладающие противоопухолевым, противовоспалительным и общетонизирующим действием. У представителей семейств болетовые, агариковые и мухоморовые гименофор расположен на нижней стороне мягкомясистых плодовых тел, имеющих хорошо различимые центральную ножку («пенек») и шляпку. Виды семейства болетовые имеют плодовые тела разной окраски с трубчатым гименофором. Почти все представители данного семейства вступают в симбиоз с корнями высших растений, образуя экзотрофную микоризу (гриб оплетает корень, оставаясь на его поверхности). Наиболее ценен белый гриб (Boletusedulis), он образует микоризу со многими лиственными и хвойными породами. Иногда характерна приуроченность к определенным типам леса и видам древесных растений: подосиновик (B. aurantiacus) - в осиновых лесах, подберезовик (B. scaber) - в березовых. Кроме того, перечисленные представители относятся к съедобным грибам – их плодовые тела используют в пищу (рис. 9. 10).  Рис. 9. 10. Белый гриб. Представители семейств агариковые (шампиньон) и мухоморовые (мухомор вонючий, бледная поганка) имеют пластинчатый гименофор. Большинство видов - сапротрофы, редко паразиты. К семейству мухоморовые принадлежат многие ядовитые грибы (рис. 9. 11 -12).  Рис. 9. 11. Мухомор.  Рис. 9. 12. Шампиньон. Класс Фрагмобазидиомицеты (Phragmobasidiomycetes). Включает типичные грибы – паразиты, не имеющие плодовых тел. Базидии разделены перегородками на четыре клетки-членика. Наиболее важное значение имеют два подкласса: головневые (Ustilaginidae) и ржавчинные (Uredinidae). Головневые - паразиты различных травянистых растений, многие из них паразитируют на зерновых культурах. Представители головневых - это узкоспециализированные паразиты, поражающие большинство органов своих растений-хозяев (почки, листья, стебли, плоды, семена, цветки и различные части соцветий). Реже они развиваются на корнях. Инфицированные части растений выглядят как обугленные или покрытые сажей – отсюда и название болезни «головня». Представители: твердая головня пшеницы (Tilletiatrutiсa) и пыльная головня пшеницы (Ustilagotrutica). Ржавчинные - паразиты различных травянистых и древесных растений. Пораженные листья и молодые стебли приобретают пятна или полосы, обычно, ржаво-бурого цвета (отсюда и название). Эти грибы имеют сложный цикл развития, который может протекать как в одном, так и в нескольких растениях (однохозяинные и разнохозяинные виды). Класс Гетеробазидиомицеты (Heterobasidiomycetes) в большинстве своем сапротрофы (на гниющей древесине), но встречаются среди них и паразиты (нередко паразитируют на плодовых телах других грибов). Представители данного класса характеризуются наличием сложной многоклеточной базидии и студенистых плодовых тел, которые в дождливую погоду выступают на сухих веточках или отмершей коре деревьев в виде студенистых подушечек разнообразной окраски. Отдел Дейтеромикоты, или несовершенные грибы (Deuteromycota) Объединяют около 30 тыс. видов. Имеют развитый многоклеточный мицелий. Размножаются, в основном, бесполым путем с помощью конидий (рис. 9. 13), половой процесс неизвестен (отсюда и название – несовершенные). 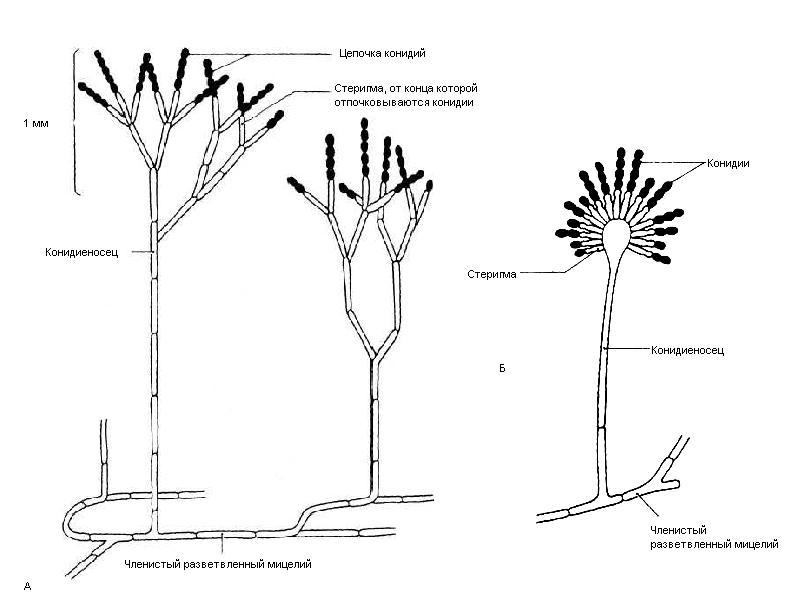 Рис. 9. 13. Бесполое размножение у двух типичных представителей Deuteromycota: А – пеницилл (Penicillium), конидиеносец имеет вид микроскопической кисточки; Б – аспергилл (Aspergillus), шаровидно вздутый на вершине конидиеносец несет радиально расходящиеся цепочки конидий. По сути, несовершенные грибы считаются формальным таксоном, т.к. они являются бесполыми формами представителей вышеназванных отделов (аскомикотов и базидиомикотов) и имеют с ними много общего в морфологии, биологии и биохимии. Представители данного отдела широко распространены в природе и обитают на различных субстратах. Среди них есть сапротрофы, которые вместе с другими грибами принимают участие в разложении органических остатков и в почвообразовательном процессе, а также паразиты. Так, представители рода фузариум (Fusarium), выделяя фузариотоксин, поражают сеянцы культурных растений и вызывают болезни растений – фузариозы. Заболевание мучнистая роса поражает смородину, крыжовник и вызвана видами мучнисторосяных грибов. Дейтеромикоты способны выделять высокотоксичные соединения (микотоксины), которые обладают и канцерогенным действием (например, афлатоксины, производимые аспергиллом паразитическим). Виды таких родов, как пеницилл (Penicillium) и аспергилл (Aspergillus), служат источниками промышленного производства ряда антибиотиков, ферментов и органических кислот. В лечебную практику постоянно вводят все новые и новые, более эффективные синтетические производные, исходным сырьем для которых по-прежнему остается природный пенициллин, в больших количествах получаемый из промышленной культуры этого гриба. Отдел лишайники – Phycomycota(Lichenes) Представители отдела чрезвычайно широко распространены в природе. Обитают от пустынь до Арктики и Антарктики. Насчитывают около 25 тыс. видов лишайников. Лишайники – это симбиотические комплексы организмов, которые образованы грибом (гетеротрофный микобионт) и водорослями или цианобактериами (автотрофный фикобионт). Гриб обеспечивает защиту водорослей от высыхания и действия крайних температур, а также снабжает их водой и минеральными солями. Водоросль снабжает гриб созданными ею органическими веществами. Вегетативное тело лишайников – слоевище, или таллом. Окраска слоевища обусловлена различными пигментами и специфическими органическими соединениями (лишайниковые кислоты). Последних около 300, и они откладываются на поверхности гиф в виде кристаллов, палочек, зернышек и придают талломам лишайников серую, сизую, зеленоватую, желтую, оранжевую, черную и др. окраску. Основу вегетативного тела лишайников (плектенхиму) образуют переплетенные членистые гифы лишайниковых грибов. Некоторые гифы имеют толстую оболочку, способную разбухать, впитывая воду. Кроме того, имеются жировые гифы, содержащие капли жира. В составе лишайников около 20 тыс. грибов, преимущественно аскомикот. Причем, грибной компонент (микобионт) каждого вида лишайника специфичен и отличается от микобионта других лишайников. Фотосинтезирующие организмы в составе лишайников представлены одноклеточными и нитчатыми зелеными водорослями (у большинства), а также цианобактериями. В отличие от микобионта один и тот же вид водоросли может быть фикобионтом нескольких видов лишайников. Всего в лишайниках встречается около 30 видов водорослей. Лишайниковые грибы неспособны к самостоятельному существованию, тогда как лишайниковые водоросли, выделенные в чистые культуры, могут самостоятельно существовать. Кроме того, установлено, что гифы гриба способны давать отростки – гаустории, которыми он внедряется в клетки водорослей и поглощает их содержимое, при этом водоросль погибает. Паразитизм гриба умеренный; погибает лишь незначительная часть водорослевых клеток. По морфологии талломов различают три основные группы лишайников: Накипные, или корковые – тело в виде корочек или накипи, связанное с субстратом всей поверхностью и практически неотделимое от него. К этой группе принадлежит до 80 % всех лишайников (рис. 9. 14). 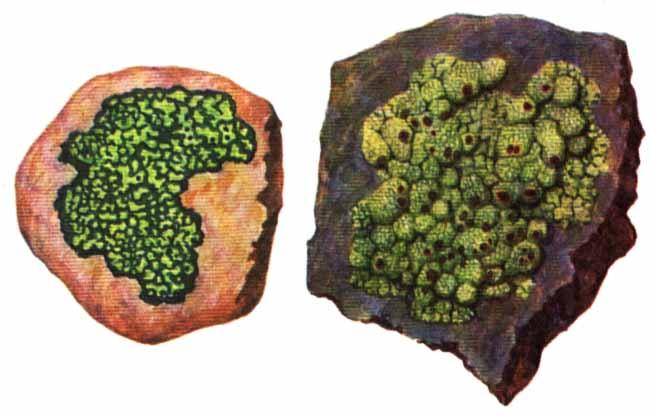 Рис. 9.14. Накипные лишайники. Листоватые – тело в виде листовидных пластинок, прикрепленных к субстрату пучками гиф и легко отделяющихся от него. У слоевища можно отличить верхнюю и нижнюю поверхности ( рис. 9. 15).  Рис. 9. 15. Листоватые лишайники. Кустистые – таллом в виде более или менее разветвленного кустика, поднимающегося с земли, или свисающего с ветвей (рис. 9. 16).  Рис. 9. 16. Кустистые лишайники. По характеру анатомического строения таллома лишайники делят на: гетеромерные и гомеомерные (рис. 9. 17). 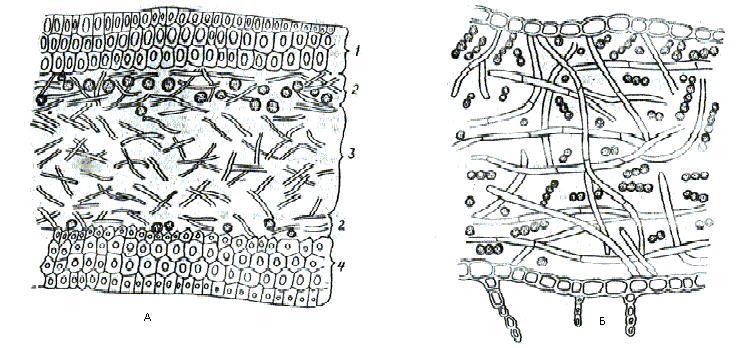 Рис. 9. 17. Строение талломов лишайников: А – гетеромерное строение: 1 – верхний коровой слой, 2 – гонидиальный (альгальный) слой, 3 – сердцевинный слой, 4 – нижний коровой слой; Б – гомеомерное строение. У гетеромерных лишайников таллом состоит из нескольких слоев. В их слоевище различают верхнюю и нижнюю кору. Последние образованы плотно сплетенными гифами микобионта. От нижней коры отходят ризины (пучки гиф), а под верхней корой размещается гонидиальный (альгальный) слой, состоящий из массы водорослей, располагающихся между переплетающимися гифами грибов. Глубже находится «сердцевина», которая образована рыхлым сплетением гиф грибов. Данную структуру имеют большинство лишайников. У гомеомерных лишайников водоросли равномерно, по всей толще, распределены среди переплетающихся гиф грибов. Такое строение свойственно накипным и небольшой части листоватых и кустистых лишайников. Размножаются лишайники преимущественно вегетативно – обломками слоевища или специальными органами – соредиями и изидиями. Отделение участков слоевища происходит механически (особенно в сухую погоду, когда они становятся хрупкими). Соредии образуются внутри водорослевого слоя и состоят из небольшого числа клеток водоросли и оплетающих их гиф гриба. Через разрыв в верхней коре слоевища соредии выпадают наружу и разносятся ветром, в благоприятных условиях они образуют новый таллом. Изидии также состоят из клеток водоросли и оплетающих их гиф гриба, но образуются в виде выростов на поверхности таллома. Каждый из компонентов лишайника способен размножаться самостоятельно. Гриб может размножаться спорами, прорастающими в мицелий. Но лишайник образуется лишь в том случае, если гифы гриба встретят на своем пути соответствующую водоросль, которая размножается делением клеток. Рост лишайников происходит очень медленно. За год их таллом нарастает у разных видов от 1 до10 мм. Продолжительность жизни таллома у отдельных видов может достигать 100 лет и более. Лишайники первыми поселяются в самых бесплодных местах суши. Отмирая, образуют перегной, на котором могут жить другие растения. По отношению к субстрату и факторам окружающей среды лишайники подразделяют на ряд экологических групп: эпигейные - растут на почве, эпифитные – на живых растениях, эпиксильные - на обработанной или гниющей древесине, эпилитные – на камнях. Основное условие поселения лишайников - длительная неподвижность субстрата. В лишайниках накапливаются полисахариды, а белков и жиров мало. Некоторые химические вещества лишайников обладают антимикробными свойствами. Кроме того, представители этого отдела используются для получения антибиотиков (кладония, пармелия, эверния и др.), ароматических веществ (некоторые лишайники синтезируют ценное эфирное масло) и фиксаторов запахов (лобария, эверния), а также красителей шерсти. Лишайники чувствительны к загрязнению воздуха (погибают при высокой концентрации двуокиси серы и других загрязнителей), поэтому используются в качестве биоиндикаторов степени загрязнения окружающей среды. |