Гилберт С. Биология развития. Т.2.doc ,БИР. Библиография Гилберт С. Биология развития в 3х т. Т. 2 Пер с англ. М. Мир, 1994. 235 с
 Скачать 19.05 Mb. Скачать 19.05 Mb.
|

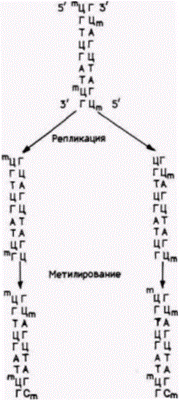
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с.__________ МЕХАНИЗМЫ ДИФФЕРЕНЦИАЛЬНОЙ ТРАНСКРИПЦИИ ГЕНОВ_________________________ 157 ная ДНК была неспособна вызвать синтез фермента в клетке-хозяине ГФРТ–. Когда ДНК получали из клона клеток, имеющих ген ГФРТ на активной Х-хромосоме, в трансформированных клетках фермент синтезировался. Годом позже было получено еще одно подтверждение того, что метилирование играет в инактивации Х-хромосомы определенную роль: тогда было обнаружено, что 5-азацитидин локально реактивирует гены на неактивной Х-хромосоме. Группа исследователей сконструировала гибридные клетки, в которых были активная Х-хромосома мыши и неактивная Х-хромосома человека (Mohandas et al., 1981). После обработки 5-азацитидином были обнаружены клоны клеток, которые синтезировали фермент ГФРТ с Х-хромосомы человека. И наконец, оказалось, что некоторые конститутивные ферменты, кодируемые Х-хромосомой (т. е. те ферменты, которые должны присутствовать в каждой клетке, а не только в специализированных), имеют на 5’-концах своих генов кластеры из ЦГ-богатых участков. В активной Х-хромосоме по крайней мере у трех генов 5'-кластеры гипометилированы, тогда как эти же кластеры в аналогичных генах на неактивной Х-хромосоме интенсивно метилированы (Wolf et al., 1982; 1984; Keith et al., 1986). Сходным образом группы ЦфГ на 3'-концах генов более метилированы в неактивной Х-хромосоме. чем в активной. Когда Х-хромосома реактивируется (естественным образом, как в ворсинках хориона или в опыте с 5-азацитидином). эти кластеры становятся гипометилированными 1 (Wolf et al., 1984). Помимо этого, когда клетки тератокарциномы дифференцируются и одна из Х-хромосом инактивируется. ЦфГ-кластеры на этих генах становятся метилированными (Lock et al., 1987). Хотя инициатор инактивации Х-хромосомы до сих пор неизвестен, весьма вероятно, что метилирование ДНК поддерживает неактивное состояние Х-хромо- 1 Большая часть изученных к настоящему времени генов, зависимых от РНК-полимеразы II, являются генами, продукты которых (например, гемоглобин, овальбумин и коллаген) характеризуют клетку определенного типа. Это вызвано тем, что такие гены производят необычно большое количество конкретных мРНК, которые могут быть легко изолированы и затем использованы как зонды для клонированных генов. Эти гены регулируются в развитии так, что их экспрессия происходит в специфических типах клеток в определенные периоды времени. Те гены, которые не регулируются в развитии (их продукты присутствуют в большинстве клеток на всех стадиях), могут контролироваться другим образом и иметь другую структуру. Для таких генов не нужны последовательности ТАТА или ЦААТ (Mellon et al., 1986; Reynolds et al., 1984). Они, по-видимому, должны иметь кластеры ЦфГ вблизи 5’-концов. Эти кластеры обычно гипометилированы и находятся в участках, гиперчувствительных к ДНКазе I (Wolf, Migeon, 1984)
сомы и передает его от одного поколения клеток к другому (Kaslow, Migeon, 1987). Если предложенная гипотеза о деметилировании справедлива, то распределение метилированных и деметилированных цитозинов должно устойчиво наследоваться на протяжении многих клеточных делений и «стираться» в половых клетках. Так в действительности и происходит. Установленный в клетках специфический характер метилирования стабилен на протяжении более 100 клеточных делений (Wigler et al., 1981; Stein et al., 1982). Предложен механизм для передачи этого распределения (Stein et al., 1982). Когда исследователи трансфицировали клетку гемиметилированной ДНК (т.е. ДНК. в которой только одна цепь метилирована), то метилирование ЦГ было обнаружено у всего клонального потомства этой клетки после многих клеточных делений. Предположили, что ЦГ-специфический метилирующий фермент использует метилированные цитидины как матрицу для метилирования цитидинов на компле- |