Гилберт С. Биология развития. Т.1.doc , БИР. Библиография Гилберт С. Биология развития в 3х т. Т. I Пер с англ. М. Мир, 1993. 228 с
 Скачать 10.9 Mb. Скачать 10.9 Mb.
|

Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.66 ГЛАВА 2
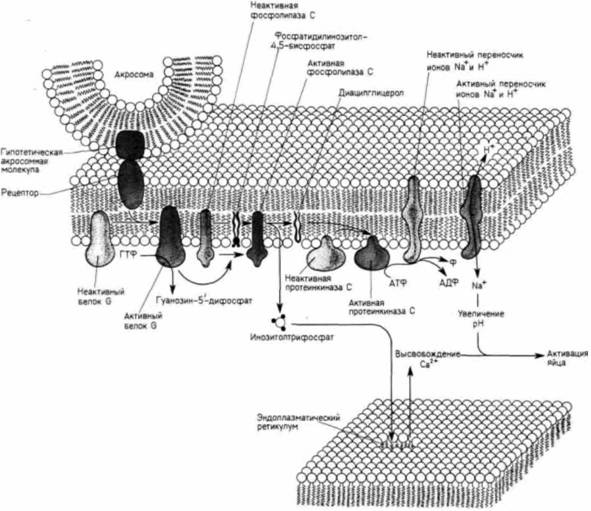
Рис. 2.35. Схема инициации ионных потоков при оплодотворении. Когда спермий прикрепляется к своему рецептору на плазматической мембране яйца, рецептор активирует белок, связывающий ГТФ (белок G), который в свою очередь активирует фосфолипазу С Фосфолипаза С расщепляет фосфатидилинозитол-4.5-бисфосфат на диацилглицерол и инозитолтрисфосфат (ИТФ). ИТФ вызывает выход Са+ из эндоплазматического ретикулума, тогда как диацилглицерол активирует протеинкиназу С. Под действием этой протеинкиназы начинает функционировать Na+/ H+-насос, в результате чего происходит обмен водородных ионов яйца на внеклеточные ионы натрия: таким образом повышается внутриклеточный pH. (Сходная схема, приводимая в гл. 20. была предложена для начала опухолевого роста.) гранул (Eisen. Reynolds, 1985). Что же происходит в течение этих 25 с. что приводит к высвобождению связанных ионов кальция? Исследования последних лет свидетельствуют о том, что прикрепление спермия к плазматической мембране яйца запускает ряд реакций, осуществляющихся с помощью заключенных в мембране ферментов. Эти ферменты синтезируют вещество, способное диффундировать в цитоплазму. Таким веществом, как было показано, является инозитол-1,4,5-трисфосфат (ИТФ), вызывающий высвобождение связанного кальция в клетках разного типа (Вeгridge,. 1985: Swann. Whitaker. 1986). В яйцах морского ежа сразу после оплодотворения синтез ИТФ возрастает (Ciapa, Whitaker. 1986). Образование ИТФ сопровождается выходом ионов кальция. Инъекции ИТФ непосредственно в яйца морского ежа и лягушки вызывают высвобождение ионов кальция и кортикальную реакцию (Whitaker, Irvine, 1984; Busa et al., 1985). Процесс выхода кальция из внутриклеточных депо при этом неотличим от реакции, наблюдаемой при нормальном оплодотворении. Кроме того, эти ИТФ-зависимые эффекты можно предотвратить, инъецируя в неоплодотворенные яйца вещества, образующие хелаты кальция (Turner et al.. 1986): это подтверждает представление о том, что ИТФ вызывает выход кальция из внутриклеточных депо. Ферментом, ответственным за синтез ИТФ, является фосфолипаза С. Этот фермент, входящий в состав плазматической мембраны, расщепляет фосфатидилинозитол-4,5-бисфосфат на диацилглицерол и ИТФ (рис. 2.35). Если ИТФ вызывает выход кальция из внутриклеточных депо, то диацилглицерол, Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с._______________ ОПЛОДОТВОРЕНИЕ: ВОЗНИКНОВЕНИЕ НОВОГО ОРГАНИЗМА 67 по-видимому, активирует другой содержащийся в мембране фермент, протеинкиназу С, которая в свою очередь активирует Na+,H+-переносчик, осуществляющий обмен ионов натрия на ноны водорода (Swann, Whitaker. 1985: Shen. Burgart. 1986). Таким образом, расщепление фосфатидилинозитол-4,5-бисфосфата может вызвать как волну высвобождения кальция, так и повышение внутриклеточного pH — два процесса, необходимые для активации яйца и его деления. Но почему прикрепление спермия к яйцу приводит к образованию ИТФ и диацилглицерола? Имеются данные, что в некоторых клетках-мишенях гормонов соединение рецептора гормона с фосфолипазой С опосредовано белком, связывающим ГТФ (Gбелок, см. Berridge. 1985). Тернер и др. (Turner et al.. 1986) показали, что то же самое происходит, по-видимому, в случае яиц морского ежа. Когда в яйцо инъецировали аналог ГТФ, стабилизирующий G-белок в активной конфигурации, кортикальные гранулы выделяли свое содержимое в отсутствие спермия. Если же в яйцо инъецировали конкурирующий ингибитор ГТФ, то вызываемый спермием нормальный процесс экзоцитоза кортикальных гранул подавлялся. Это подавляющее действие можно было предотвратить, инъецируя в неоплодотворенное яйцо ИТФ. Схема начальных биохимических событий при активации яйца представлена на рис. 2.32 и 2.35. Прикрепление спермия к неидентифицированному рецептору на плазматической мембране яйца изменяет конформацию этого рецептора таким образом, что он активирует G-белок. G-белок в свою очередь активирует фосфолипазу С, которая расщепляет фосфатидилинозитол-4,5-бисфосфат на два внутриклеточных посредника, диацилглицерол и инозитолтрисфосфат. Диацилглицерол активирует протеинкиназу С; последняя фосфорилирует белок-переносчик Na+ и H+, тем самым делая его способным функционировать (при стимуляции ионами кальция), что приводит к повышению внутриклеточного pH. Поскольку ИТФ вызывает выход кальция из эндоплазматического ретикулума, диффузия ИТФ приводит к распространению волны выхода кальция по всему кортикальному слою яйца. Таким образом, прикрепление спермия к плазматической мембране вызывает определенные изменения внутри яйца. Преобразования цитоплазмы яйцаОплодотворение может также вызывать радикальные перемещения компонентов цитоплазмы. Эти перемещения часто играют решающую роль при последующем развитии в процессах клеточной дифференцировки. Как мы увидим в гл. 7 и 8, цитоплазма яйца нередко содержит морфогенетические детерминанты, которые сегрегируются в процессе дробления и попадают в определенные клетки. Эти детерминанты в конечном счете приводят к активации или репрессии специфических генов: в результате клетки, содержащие такие детерминанты, приобретают определенные свойства. Правильное пространственное распределение морфогенетических детерминантов является решающим условием нормального развития. У некоторых видов благодаря присутствию пигментных гранул можно проследить перемещение таких детерминантов в соответствующую область. Одним из примеров может служить яйцо асцидии Styela partita(Conklin, 1905). Неоплодотворенное яйцо этого животного представлено на рис. 2.36.А. Серая цитоплазма центральной части яйца окружена кортикальным слоем, содержащим желтые липидные включения. Во время мейоза разрушение ядерной оболочки освобождает светлый ядерный материал, который накапливается в анимальном (верхнем) полушарии яйца. В течение 5 мин после проникновения спермия внутренняя светлая и кортикальная желтая цитоплазма перетекает в вегетативное (нижнее) полушарие яйца. Когда затем мужской пронуклеус мигрирует от вегетативного полюса к экватору по будущей задней стороне зародыша, липидные включения перемещаются вместе с ним. В результате этой миграции возникает желтый серп, занимающий пространство от вегетативного полюса до экватора (рис. 3.36, Б—Г), и желтая цитоплазма оказывается в области, где позднее у личинки образуются мышечные клетки. Перемещения цитоплазмы наблюдаются и в ооцитах амфибий. У многих видов бесхвостых амфибий единственный спермий может проникнуть в любом месте анимального полушария; после этого события распределение цитоплазматических компонентов яйца изменяется. Наиболее заметны изменения в пигментированном поверхностном слое цитоплазмы. Первоначально слой темного меланинового пигмента покрывает все анимальное полушарие. После проникновения спермия поверхностный слой цитоплазмы смещается примерно на 30о по отношению к внутренней цитоплазме (Manes, Elinson. 1980: Vincent et al.. 1986), в результате чего у некоторых бесхвостых амфибий (например, у лягушек рода Rann) вблизи экватора против места проникновения спермия появляется серповидная слабо пигментированная область (рис. 2.37) (Roux. 1887, Ancel. Vintenberger, 1948). Эту слабо пигментированную область назвали серым серпом. Как мы увидим в последующих главах, серый серп соответствует области, в ко- |