Гилберт С. Биология развития. Т.1.doc , БИР. Библиография Гилберт С. Биология развития в 3х т. Т. I Пер с англ. М. Мир, 1993. 228 с
 Скачать 10.9 Mb. Скачать 10.9 Mb.
|

Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.100
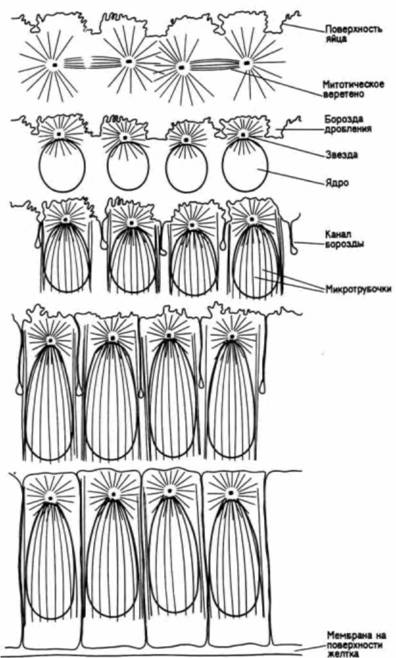
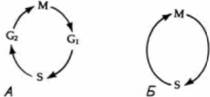
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.______ ДРОБЛЕНИЕ: СОЗДАНИЕ МНОГОКЛЕТОЧНОСТИ 101 крипции (начинающейся приблизительно на одиннадцатом цикле) сильно возрастает. Замедление ядерных делений и ускорение синтеза РНК у дрозофилы напоминают ситуацию, наблюдающуюся у зародышей лягушки и морского ежа. Контроль за замедлением делений (у зародышей Xenopus, морского ежа и дрозофилы), по-видимому, осуществляется через соотношение между ядром и цитоплазмой (Newport, Kirschner. I982a; Edgar et al., 1986). Эдгар и его соавторы в цитированной работе сравнили раннее развитие зародышей дрозофилы дикого типа с развитием гаплоидных мутантов. Гаплоидные зародыши дрозофилы при каждом клеточном делении содержат вдвое меньше хроматина, чем зародыши дикого типа. Следовательно, гаплоидный зародыш на восьмом клеточном цикле имеет такое же количество хроматина, что и зародыш дикого типа на седьмом цикле. Эти исследователи установили, что тогда как зародыши дикого типа формировали клеточную бластодерму во время 14-го цикла, гаплоидные зародыши для достижения того же результата должны были осуществить еще одно, дополнительное 15-е деление. Более того, продолжительность 11 —14 циклов у зародышей дикого типа соответствовала продолжительности 12—15 циклов у гаплоидных зародышей. Таким образом, ход развития гаплоидных зародышей сходен с ходом развития зародышей дикого типа, отличаясь лишь отставанием на одно клеточное деление. Если это отставание обусловлено тем, что ядерно-плазменное отношение у гаплоидных мутантов на каждом данном цикле вдвое меньше, чем у зародышей дикого типа, то можно было бы ускорить образование клеточной бластодермы, отшнуровывая (наложением лигатуры) часть цитоплазмы, чтобы гаплоидные ядра делились в меньшем ее объеме. Когда такой опыт был проведен, образование клеточной бластодермы у гаплоидных зародышей ускорилось. Завершающее деление клеток бластодермы, свидетельствующее об окончании периода дробления, у дрозофилы происходит тогда, когда на одно ядро приходится 61 мкм2 цитоплазмы У Xenopus сходное замедление митотических делений наблюдается после 12-го деления. И здесь деления также впоследствии становятся асинхронными. Момент перехода к средней бластуле тоже связан с ядерно-плазменным отношением. Гаплоидные зародыши Xenopuslaevis осуществляют этот переход после 13-го митотического цикла, тогда как тетраплоидные зародыши (содержащие вдвое больше хроматина на клетку, чем зародыши дикого типа) приступают к переходу после 11-го деления (Newport, Kirschner, 1982a, b). Как у дрозофилы, так и у Xenopus laevis транскрипция и подвижность клеток могут быть инициированы преждевременно путем искусственного удлинения клеточного цикла. При задержке клеточных делений циклогексимидом (ингибитор белкового синтеза) переход к средней бластуле у Xenopus laevis индуцируется рано, а у дрозофилы происходит вспышка транскрипции (Edgar et al.. 1986; Kimelman et al.. 1987). МЕХАНИЗМЫ ДРОБЛЕНИЯИнициация дробленияКлеточный цикл соматических клеток по функциональному признаку делится на четыре стадии (или фазы). За митозом (М) следует пресинтетический период (G1), после которого происходит синтез ДНК (S). Затем наступает премитотический цикл (G2), за которым следует митоз (рис. 3.39.А). Наступление этих фаз регулируется факторами роста, которые будут подробно рассмотрены в гл. 20. Клеточный цикл бластомеров на ранних стадиях дробления может быть, однако, гораздо более простым. Бластомеры у морского ежа на этих стадиях не имеют фазы G1; репликация ДНК осуществляется в телофазе предыдущего митоза (Hinegardner et al., 1964). На тех же стадиях у Xenopuslaevisи дрозофилы отсутствуют две фазы, G1 и G2. (У Xenopuslaevisэти фазы появляются в клеточном цикле при переходе к средней бластуле после 12-го деления, а у дрозофилы — после 14-го. В обоих случаях это изменение цикла сочетается с увеличением клеточной подвижности и транскрипции РНК.) В течение первых 12 делений клетки Xenopusделятся синхронно и клеточный цикл бывает двухфазным: S-фаза сменяется М-фазой и M фаза —S-фазой (рис. 3.39, Б. Laskey et al., 1977; Newport, Kirschner, 1982a). Факторы, регулирующие этот двухфазный цикл, локализованы в цитоплазме. В норме преобразования растущих ооцитов Xenopuslaevisостанавливаются на стадии профазы первого деления мейоза. Такие ооциты неспособны к делению. Когда в них пересаживают ядра делящихся клеток, эти ядра перестают делиться. Нормальные ооциты, подвергнутые стимулирующему воздействию прогестерона, возоб-
|