Билет 1 биологическое окисление
 Скачать 1.13 Mb. Скачать 1.13 Mb.
|

|
Билет 1 1. БИОЛОГИЧЕСКОЕ ОКИСЛЕНИЕ - окислительно-восстановительные реакции, протекающие в организмах; осн. источник их энергии. катализируется –оксидоредуктаз. При анаэробном Б. о. акцептором водорода являются низкомолекулярные соединения (НАД или НАДФ). При аэробном Б. о,) акцептором водорода служит кислород; ткани при этом поглощают кислород и выделяют CO2 и Н2О. В результате Б. о. освобождённая энергия накапливается в дыхательной цепи в виде химич. связей макроэргич. соединений. Энергия освобождается в процессе ферментативного окисления метаболитов специфическими дегидрогеназами. В реакциях дегидрирования электроны и протоны переходят от органических субстратов на коферменты NAD- и FAD-зависимых дегидрогеназ. Восстановление молекулы О2 происходит в результате переноса 4 электронов. При каждом присоединении к кислороду 2 электронов, поступающих к нему по цепи переносчиков, из матрикса поглощаются 2 протона, в результате чего образуется молекула Н2О. процесс переноса электронов по ЦПЭ и синтез АТФ энергетически сопряжены. Первый процесс - перенос электронов от восстановленных коферментов NADH и FADH2 через ЦПЭ на кислород - экзергонический. Например: NADH + Н+ +1/2 O2 → NAD+ + H2O + 52 ккал/моль(≈220 кДж/моль). (1) Второй процесс - фосфорилирование АДФ, или синтез АТФ, - эндергонический: АДФ + Н3РО4+7,3 ккал/моль (30,5 кДж/моль) = АТФ + Н2О. (2) Синтез АТФ из АДФ и Н3РО4 за счёт энергии переноса электронов по ЦПЭ называют окислительным фосфорилированием. 2.субстратное S---OP3H2+АДФ→АТФ+S (ФЕРМЕНТ-КИНАЗА) 3.ФОТОФОСФОРИЛИРОВАНИЕ H2O→NADH+H++O2 (hv-квант света) NADH+H++АДФ+P→АТФ+NAD+ 2.Синтези распад гликогена: Гликоген синтезируется в период пищеварения (через 1-2 ч после приёма углеводной пищи). синтез гликогена из глюкозы, как и любой анаболический процесс, является эндергоническим, т.е. требующим затрат энергии. синтез гликогена осуществляется путём удлинения уже имеющейся молекулы полисахарида"праймер". Распад гликогена происходят в ответ на повышение потребности организма в глюкозе. Гликоген печени распадается в интервалах между приёмами пищи, кроме того, этот процесс в печени и мышцах ускоряется во время физической работы. Распад гликогена происходит путём последовательного отщепления остатков глюкозы в виде глюкозо-1-фосфата. Гликозидная связь расщепляется фермент гликогенфосфорилазой. чем больше концов имеет молекула гликогена, тем больше молекул гликогенфосфорилазы могут действовать одновременно. 3. Фосфолипи́ды — сложные липиды, сложные эфиры многоатомных спиртов и высших жирных кислот. Содержат остаток фосфорной кислоты и соединенную с ней добавочную группу атомов различной химической природы. 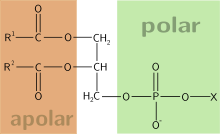
глицерофосфолипиды (глицерофосфатиды) — содержат остаток глицерина фосфатидилхолин (лецитин) фосфатидилэтаноламин (кефалин) фосфатидилсерин кардиолипин плазмалоген(этаноламиновый плазмологен) фосфосфинголипиды — содержат остаток сфингозина сфингомиелины фосфоинозитиды — содержат остаток инозитола фосфатидилинозитол Фосфолипиды — амфифильные вещества. Главная особенность фосфолипидов состоит в том, что «головка» у них гидрофильна, а «хвосты» гидрофобны. образовывать бислой — двойной слой фосфолипидных молекул, где гидрофильные головы с обеих сторон соприкасаются с водой, а гидрофобные хвосты упрятаны внутрь бислоя. Ф.-Главный липидный компонент клеточных мембран. Они служат источником фосфорной к-ты, необх для жизни человека. В кл мембранах обеспечивают текучие и пластические свойства мембран клеток и кл органоидов. входят в состав липопротеидов кл мембран. Ф. участвуют в транспорте жирных кислот и холестерина. Ф. замедляют синтез коллагена и повышают активность коллагеназы . Сфинголипиды Аминоспирт сфингозин, состоящий из 18 атомов углерода, содержит гидроксильные группы и аминогруппу. Продукт взаимодействия сфингозина и жирной кислоты называют "церамид").. Церамиды отличаются радикалами жирных кислот, входящих в их состав. Существует 3 основных типа сфинголипидов: Церамиды Сфингомиелины Гликосфинголипиды Биол. роль сфинголипидов разнообразна. Известно, что они участвуют в формировании мембранных структур аксонов, синапсов и др. клеток нервной ткани, опосредуют в организме механизмы узнавания, рецепторные взаимодействия, межклеточные контакты и др. жизненно важные процессы. Глицерофосфолипиды Структурная основа - глицерол. Глицерофосфолипиды молекулы, в которых две жирные кислоты связаны сложноэфирной связью с глицеролом в первой и второй позициях; в третьей позиции находится остаток фосфорной кислоты Фосфатидная кислота в свободном состоянии в организме содержится в небольшом количестве), но является промежуточным продуктом на пути синтеза как три-ацилглицеролов, так и глицерофосфолипидов. У глицерофосфолипидов во второй позиции находятся преимущественно полиеновые кислоты Фосфатидилэтаноламины. являются главными липидными компонентами мембранклеток. Фосфатидилсерины они участвуют в синтезе фосфатидилэтаноламинов. Фосфатидилинозитолы. Они обнаружены у животных, растений и микроорганизмов. В животноморганизме найдены в мозге, печени и легких. Гликолипиды Главной формой гликолипидов в животных тканяхявляются гликосфинголипиды. кот. содержат церамид, состоящий из спирта сфингозина и остатка жирной кислоты, и один или несколько остатков сахаров. Простейшими гликосфинголипидами являются галактозилцерамиды и глюкозилцерамиды. Галактозилцерамиды – главные сфинголипиды мозга и других нервных тканей, но в небольших количествах встречаются и во многих других тканях Глюкозилцерамиды – простые гликосфинголипиды, представлены в тканях, отличных от нервной, причем главным образом глюкозил-церамидами. В небольших количествах они имеются в ткани мозга. В отличие от галактозилцерамидов у них вместо остатка галактозы имеется остаток глюкозы. Более сложными гликосфинголипидами являются ганглиозиды, образующиеся из гликозилцерамидов. Ганглиозиды в больших количествах находятся в нервной ткани выполняют рецепторные и другие функции. Билет №2 1 (9) 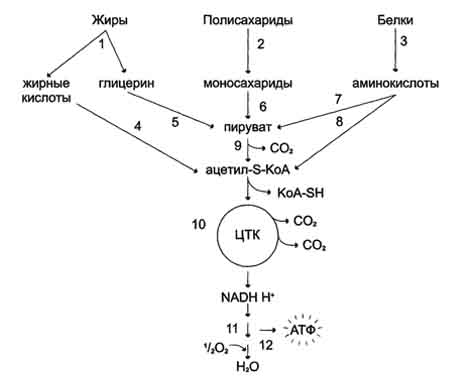 Специфический путь катаболизма-это совокупность превращений, свойственных только 1 соединению. Из глюкозы при гликолизе образуется ПВК. Из жирных кислот при бета-окислении обр-ся Ко-А. Аминокислоты образуют ПВК. В результате специфических путей катаболизма все в-ва превращаются в пвк и ацетил-КоА, которые затем направляются в общий путь катаболизма, включающий в себя процесс декарбоксилирования пировиноградной кислоты и цикл трикарбоновых кислот. который является источником основной массы первичных доноров Н для дыхательной цепи. Окислительное декарбоксилирование пвк: H3CC(О) COOH+ HS-KoA + НАД+→Н3С-C(O)- S- KoA + НАДН + Н+ + СО2 Пируват Ацетил-КоА В результате этой реакции образуются ацетил-КоА, восстановленная форма (НАДН + Н+) и диоксид углерода. 2(16). Углево́ды — орг в-ва, содержащие карбонильную группу и несколько гидроксильных групп.. Углеводы участвуют в построении различных опорных структур. целлюлоза - осн структурным компонентом кл стенок растений, хитин выполняет аналогичную функцию у грибов, обеспечивает жёсткость экзоскелета членистоногих.Пластическая функция. Углеводы входят в состав сложных молекул участвуют в построении АТФ, ДНК и РНК. Энергетическая функция. Углеводы служат источником энергии: при окислении 1 грамма углеводов выделяются 4,1 ккал. Углеводы выступают в качестве запасных питательных веществ: гликоген у животных, крахмал— у растени. Олигосахариды входят в состав воспринимающей части многих клеточных рецепторов. Моносахариды-производные многоатомных спиртов, содержащие карбонильную группу. подразделяют на альдозы и кетозы.Альдозы содержат функциональную альдегидную группу -НС=О, кетозы содержат кетонную группу >С=О. Расположение Н- и ОН-групп относительно пятого углеродного атома определяет принадлежность глюкозы к D- или L-ряду. В организме чел моносахариды находятся в D-конфигурации, α- и β-формы, обозначают определённую конформацию Н- и ОН-групп относительно С1. У α-D-глюкозы ОН-группа располагается ниже плоскости кольца, а у β-D-глюкозы, наоборот, над плоскостью кольца. Глюкозаявляется альдогексозой. существует линейнаяи циклическаяформах. Циклическая форма глюкозы,, обусловливает химические свойства глюкозы. глюкоза имеет 4 асимметричных углеродных атома, Возможно образование 16 стереоизомеров, наиболее важные D- и L-глюкоза. В организме млекопитающих моносахариды находятся в D-конфигурации Фруктозаявляется кетогексозой. Фруктоза существует в циклической форме, образуя α- и β-аномеры Олигосахариды Олигосахариды содержат от двух до десяти остатков моносахаридов, соединённых гликозидной связью. Дисахариды - наиболее распространённые олигомерные углеводы(как сахароза, лактоза и мальтоза) Полисахариды можно разделить на гомополисахариды (все мономеры идентичны) и гетерополисахариды (мономеры различны). В зависимости от выполняемых ими функций полисахариды можно разделить на 3 основные группы: резервные полисахариды, выполняющие энергетическую функцию, структурные полисахариды, обеспечивающие клеткам и органам механическую прочность и полисахариды, входящие в состав межклеточного матрикса, принимают участие в образовании тканей 3Реакции синтеза холестерола происходят в цитозоле клеток. 1. Образование мевалоната. Две молекулы ацетил-КоА конденсируются ферментом тиолазой с образованием ацетоацетил-КоА.. Фермент щдроксиметилглутарил-КоА-синтаза присоединяет третий ацетильный остаток с образованием ГМГ-КоА. Следующая реакция, катализируемая ГМГ-КоА-редуктазой, является регуляторной в метаболическом пути синтеза холестерола. восстановление ГМГ-КоА до мевалоната с использованием 2 молекул NADPH. Фермент ГМГ-КоА-редуктаза 2. Образование сквалена На втором этапе синтеза мевалонат превращается в пятиуглеродную изопреноидную структуру, содержащую изопентенилпирофосфат- геранилпирофосфат – фарнезилпирофосфата. Две молекулы фарнезилпирофосфата конденсируются с образованием сквалена 3. Образование холестерола сквален превращается в ланостерол. Далее происходит 20 последовательных реакций, превращающих ланостерол в холестерол. На последних этапах синтеза от ланостерола отделяется 3 атома углерода. 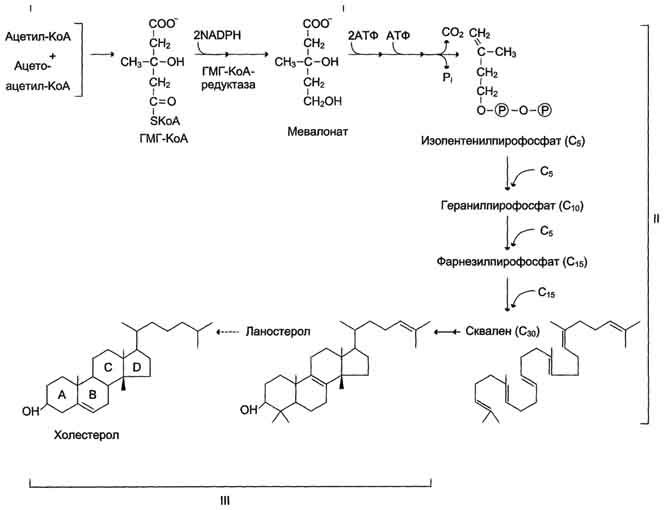 Регуляция синтеза холестерола. Регуляция активности ГМГ-КоА-редуктазы в печени. Холестерол и жёлчные кислоты снижают скорость транскрипции и, таким образом, синтез фермента. Инсулин стимулирует дефосфорилирование, а глюкагон - фосфорилирование ГМГ-КоА-редуктазы. Инсулин активирует 2 фосфатазы: киназы ГМГ-КоА-редуктазы* и фосфатазу, дефосфорилирующую непосредственно ГМГ-КоА-редуктазу. Глюкагон стимулирует фосфорилирование и инактивацию 2 фосфатаз и фосфорилирование и активацию киназы ГМГ-КоА-редуктазы. В абсорбтивный период синтез холестерола увеличивается. увеличивается и доступность исходного субстрата для синтеза холестерола - ацетил-КоА . В результате синтез холестерола в постабсорбтивном периоде и при голодании ингибируется. холестерол снижает скорость транскрипции гена ГМГ-КоА-редуктазы, подавляя таким образом собственный синтез. В печени активно идёт синтез жёлчных кислот из холестерола, поэтому и жёлчные кислоты подавляют активность гена ГМГ-КоА-редуктазы). Билет №3 1.(11 процесс окислительного декарбоксилирования пирувата катализирует пируватдегидрогеназный комплекс. В (ПДК) входят 3 фермента: пируватдекарбоксилаза (Е1) – кофермент - ТДФ, дигидролипоилтрансацетилаза (Е2) – кф липоевая кислота и коа- и дигидролипоилдегидрогеназа (Е3),-FAD, NAD+ .в состав комплекса входят регуляторные субъединицы: протеинкиназа и фосфопротеинфосфатаза 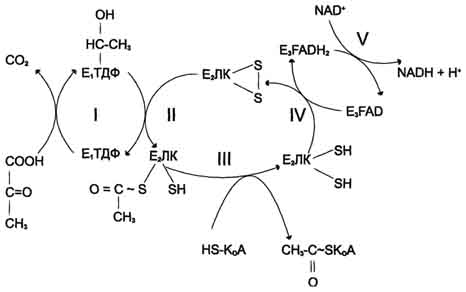 Е1 катализирует декарбоксилирование пирувата и перенос С2-фрагмента на ТДФ; II - Е2 катализирует окисление гидроксиэтильной группы и перенос С2-фрагмента на липоевую кислоту (ЛК); III - ацетилированная дигидролипоилтрансацетилаза взаимодействует с КоА с образованием восстановленной формы липоевой кислоты и ацетил-КоА; IV - окисленная форма трансацетилазы регенерируется при участии E3;V - окисленная форма Е3 востанавливается при участии NAD+ 2(20) Переваривание углеводов начинается в ротовой полости. пища измельчается при пережёвывании, смачиваясь слюной, где присутствует гидролитический фермент α-амилаза (α-1,4-гликозидаза), расщепляющая в крахмале α-1,4-гликозидные связи. амилаза слюны не расщепляет α- 1,6-гликозидные связи. крахмал переваривается частично с образованием крупных фрагментов - декстринов и небольшого количества мальтозы. Желудочный сок не содержит ферментов, расщепляющих углеводы. происходит незначительный кислотный гидролиз гликозидных связей.В тонком кишечнике в разных его отделах под действием гидролитических ферментов. В двенадцатиперстной кишке поступает панкреатическая α-амилаза. Этот фермент гидролизует α-1,4-гликозидные связи в крахмале и декстринах.Продукты переваривания крахмала – мальтоза. Панкреатическая α-амилаза не расщепляет α-1,6-гликозидные связи в крахмале. Целлюлоза проходит через кишечник неизменённой. придавая пище дополнительный объём и положительно влияя на процесс переваривания и стимулируя перистальтику кишечника.. Сахаразо-изомальтазный комплекс прикрепляется к мембране микроворсинок кишечника с помощью гидрофобного (трансмембранного) домена, образованного N-концевой частью полипептида. Каталитический центр выступает в просвет кишечника. гидролизует сахарозу и изомальтозу, расщепляя α-1,2- и α-1,6-гликозидные связи. Гликоамилазный комплекс катализирует гидролиз α-1,4-связи между глюкозными остатками в олигосахаридах, действуя с восстанавливающего конца. Комплекс расщепляет также связи в мальтозе. β-Гликозидазный комплекс (лактаза) расщепляет β-1,4-гликозидные связи между галактозой и глюкозой в лактозе. активность лактазы у плода особенно повышена в поздние сроки беременности и сохраняется до 5-7-летнего возраста. Затем активность фермента снижается. Транспорт моносахаридов в клетки слизистой оболочки кишечника может осуществлятьсяпутём облегчённой диффузии и активного транспорта. В случае активного транспорта глюкоза и Na+ проходят через мембраны с люминальной стороны, связываясь с разными участками белка-переносчика. При этом Na+ поступает в клетку по градиенту концентрации, и одновременно глюкоза транспортируется против градиента концентрации. чем больше градиент Na+, тем больше поступление глюкозы в энтероциты. Если концентрация Na+ во внеклеточной жидкости уменьшается, транспорт глюкозы снижается. Глюкозные транспортёры (ГЛЮТ) ГЛЮТ-1 обеспечивает стабильный поток глюкозы в мозг;ГЛЮТ-2 при его участии глюкоза переходит в кровь из энтероцитов и печени, участвует в транспорте глюкозы в β-клетки поджелудочной железы;ГЛЮТ-3 обеспечивает постоянный приток глюкозы к клеткам нервной и других тканей;ГЛЮТ-4 - главный переносчик глюкозы в клетки мышц и жировой ткани;ГЛЮТ-5 встречается в клетках тонкого кишечника. В основе патологии переваривания и всасывания углеводов: дефекты ферментов, участвующих в гидролизе углеводов в кишечнике; нарушение всасывания продуктов переваривания углеводов в клетки слизистой оболочки кишечника. возникает осмотическая диарея, спазмы, боли,метеоризм. 3(61) . Х. транспортируется кровью только в составе ЛПНП и ЛПВП. ЛП обеспечивают поступление в ткани экзогенного х., определяют потоки х. между органами и выведение избытка х. из организма. ЛПНП - основная транспортная форма х., в которой он доставляется в ткани. Около 70% х. и его эфиров в крови находится в составе ЛПНП. Из крови ЛПНП поступают в печень (до 75%) и другие ткани, которые имеют на своей поверхности рецепторы ЛПНП. вып. 2 осн.ф-и: поставляют апопротеины другим ЛП в крови и участвуют в "обратном транспорте х.". ЛПВП синтезируются в печени и в небольшом количестве в тонком кишечнике. В печени в ЛПВП включаются апопротеины А, Е, С-II, фермент ЛХАТ. В крови апоС-II и апоЕ переносятся с ЛПВП на ХМ и ЛПОНП. Для переноса х. На поверхности ЛПВП находится фермент ЛХАТ, кот превращает х., имеющий гидроксильную группу, выступающую на поверхность липопротеинов или мембран клеток, в эфиры холестерола. Гидрофобная молекула, эфира холестерола перемещается внутрь ЛПВП. образом, часть эфиров холестерола переносится на ЛПОНП, ЛППП, а ЛПВП3 за счёт накопления триацилглицеролов увеличиваются и превращаются в ЛПВП2. ЛПОНП под действием ЛП-липазы превращаются сначала в ЛППП, а затем в ЛПНП. ЛПНП и ЛППП захватываются клетками через рецепторы ЛПНП. Липопротеины различаются и по участию в атерогенезе. Атерогенность липопротеинов частично зависит от размера частиц. Самые мелкие липопротеины, такие как ЛПВП, легко проникают в стенку сосуда, но также легко ее покидают, не вызывая атеросклероз. ЛПНП – наиболее атерогенные липопротеины крови. достаточно малы, чтобы проникать в стенку сосуда, и легко задерживаются в сосудистой стенке. Концентрация холестерола в крови взрослых людей составляет 200±50 мг/дл (5,2±1,2 ммоль/л) и, как правило, увеличивается с возрастом. Превышение нормальной концентрации холестерола в крови называют гиперхолестеролемией. развивается вследствие избыточного поступления холестерола с пищей. Правильное питание в течение всей жизни - важнейший фактор профилактики гаперхолестеролемии. Доказана корреляция между увеличением концентрации холестерола в плазме крови и смертностью от заболеваний ССС - инфаркта миокарда и инсульта, развивающихся в результате атеросклероза. Важным лечи проф фактором, снижающим риск развития гиперхолестеролемии и атеросклероза, является гипокалорийная и гипохолестериновая диета. Поступление холестерола с пищей не должно превышать 300 мг/сут Растительная пища должна составлять основу рациона. К лечебным и профилактическим мерпм относят обогащение пищи полиеновыми жирными кислотами семейства ω-3. Антиоксиданты и Витамины С, Е, А поддерживают нормальную структуру липидов ЛПНП и их метаболизм. Наиболее эффективные препараты, применяемые при лечении атеросклероза, - ингибиторы ГМГ-КоА-редуктазы. в печени эффективно ингибируют регуляторный фермент биосинтеза холестерола. фибраты - ускоряют катаболизм ЛПОНП, активируя ЛП-липазу. |