Ответы на билеты по биологии. Билет1 Закономерности существования клетки во времени. Жизненный цикл клетки, его варианты. Основное содержание и значение периодов жизненного цикла клетки
 Скачать 2.17 Mb. Скачать 2.17 Mb.
|

|
2.2. Упорядоченность биологических систем и энергия 2.2.1. Упорядоченность биологических систем обусловлена выделением клеткой тепловой энергии, подобноТ-машинам, скорее холодильникам, создающим местный порядок и неравновесия за счет большего уравновешивания, окруженияисточников «более свободной»(см.2.4.1) энергии, первично – света 2- Солнца Фотосинтезирующими, для синтеза органических соединений (включая световые и темновые стадии,1 У.Е), их 3- Химическая энергия переходит от растений к животным, при обратной реакции питания-дыхания, 4. Клетки получают энергию в результате окисления биомолекул, 5- Распада, в результате последовательных ферментативных реакций (2.3, требуя катализа – уменьшения барьеров, энергии активации ферментами), запасая путем сопряжения (2.3.5, 2.4) 6- Часть энергии окисления в образовании АТР, «энерговалюте» (ЛИПМАН 1899-1953-86), а ее использование - 7. Гидролиз АТР обеспечивает упорядоченность в клетке (2.3, 2.4.2), в т.ч.синтез и транспорт молекул и движение клетки. Заключение 2.3. Питательные вещества и источники энергии клетки 1. Молекулы питательных веществ, расщепляясь в три этапа, образуют АТР (после гидролиза на мономеры и расщепления тех до С2-3-единиц, общих жирам, АК и углеводам), на этапе 2 - в процессе гликолиза (С6=2С3, брожения) образуемый и без кислорода(за счет ред-окс - большей выгоды СООН), но следующий этап 3. Окислительный катаболизм поставляет значительно больше энергии,- 4. Центральным процессом метаболизма является цикл лимонной кислоты (КРЕБС Ханс, 1900-53-81, СЕНТ-ДЬЁРДЬИ 1893-1937-86) - окисление ацетата (через соединение С-Н +С=О в С-С-ОН) до СО2 и Н-переносчиков, у эукариот – в митохондриях (гл.7), после 5.При окислительном фосфорилированииперенос электронов (Н) к кислороду приводит к образованию АТР и воды, посредством хемиосмотического сопряжения (ФОФ+НОН=Н+Ф-) с транспортом Н+, мембранным потенциалом(гл.7) 6. Аминокислоты и нуклеотиды принимают участие в круговороте азота (фиксируемого прокариотами и при высокой Т, молнии, и выводимого с мочевиной,аммиаком и др. Аналогично циклам С+-4иN-3/+5важны переходы S-2/+6, возможны и Г-1/+7) Заключение 2.4. Биосинтез и создание упорядоченности (транспорт внутрь и наружу связан с ката- и анаболизмом,разделением реакций и функций) 1.Возможность протекания реакции определяется величиной изменения свободнойэнергии (Гиббса или Гельмгольца, выражающей и энтропию, ее рост - 2-й закон ТД), возможны только уменьшающие ее, поэтому другие, как 2. Реакции биосинтеза (типа А-Н+В-ОН= АВ+НОН, обратно гидролизу, 5) часто сопряжены с гидролизом АТР (энергией отталкивания Ф-,аналогично отнятию воды Р2О5,с выделением ок.30 кДж/моль, для выгоды в сумме их энергий, общего изменения свободной, 1), фосфорилированием, аналогично 3. Коферменты участвуют в переносе специфических химических групп (из-за сложности получаясь нами извитаминов, см.НПХ,Б). Как переносчики ацетата (Ац-КоА) или Н (типа НАДН и ФАДН2, 2.3.4) в ОВ, ред-окс-реакциях, где при распаде образуются, а 4. Для биосинтеза необходимы восстановительные эквиваленты, чаще НАДФН. 5. Синтез био полимеров осуществляется путем повторения элементарных реакций дегидратации (за счет АТФ, фосфорилирования предшественников), обратно их гидролизу (2.3.1). Заключение 2.5. Координация катаболизма и биосинтеза- анаболизма, составляющих 1. Метаболизм - организуемый и регулируемый процесс, где тысячи реакций, в т.ч. конкурирующих, отражаемых картах («джунглях» безточных координат степени окисления-Сп) как 2- Метаболические пути регулируются изменениями ферментативной активности, при изменении условий, по принципу обратной связи (-ОС, например, конечным продуктом – первого фермента, при +ОС возможныавтокатализ иколебания), 3. Катаболические реакции могут обращаться при поглощении энергии (2.4.1), как распад и синтез глюкозы – гликолиз и глюконеогенез, регулируемые фосфофруктокиназой, активируемой АМФ, АДФ и ингибируемые – АТФ, ЖК и цитратом, обратно – бисфосфатазой. На большее время 4. Ферменты могут переходить в активное или неактивное состояния путем ковалентных модификаций, де-фосфорилирования – фосфатазами и протеинкиназами (ПК), 2). Полигенное наследование. Группы сцепления. Кроссинговер как механизм нарушения сцепления генов. Значение генетических рекомбинаций для человека.Тип наследования признаков, обусловленных действием многих генов, каждый из которых оказывает лишь слабое действие. Фенотипически проявление полигенно обусловленного признака зависит от условий внешней среды. У потомков наблюдается непрерывный ряд вариаций количественного проявления подобного признака, а не появление четко различающихся по фенотипу классов. В ряде случаев при блокировании отдельного гена признак не проявляется вообще, несмотря на его полигенную обусловленность. Это свидетельствует о пороговом проявлении признака. Группы сцепления Гены, находящиеся в одной хромосоме и наследующиеся сцепленно, составляют группу сцепления. Количество групп сцепления каждого вида должно соответствовать числу пар хромосом. Кроссинговер возникает со вполне определенной частотой для каждой пары генов, расположенных в одной группе сцепления. Причем, чем ближе в хромосоме расположены гены друг у другу, тем она выше. На основании анализа частоты кроссинговера между генами можно вычислить расстояние между генами и, таким образом, определить их локализацию в хромосоме План расположения генов в хромосоме называется картой хромосомы. Хромосомные карты построены для животных ряда видов, однако для собаки их нет, так как для их составления необходимы фундаментальные исследования генетики вида и большой статистический материал, получение которого у собак затруднительно. Для кошки установлено положение на хромосомах приблизительно для 50 генов (Бородин, 1995). На основании анализа результатов многочисленных экспериментов с мухой дрозофилой Т. Морган сформулировал свою хромосомную теорию наследственности, сущность которой заключается в следующем: 1. Материальные носители наследственности — гены — находятся в хромосомах, располагаются в них линейно на определенном расстоянии друг от друга. 2. Гены, расположенные в одной хромосоме, наследуются сцепленно. Число групп сцепления соответствует гаплоидному числу хромосом. 3. Признаки, гены которых находятся в одной хромосоме, наследуются сцепленно. 4. В потомстве гетерозиготных родителей новое сочетание генов, расположенных в одной паре хромосом, может возникнуть в результате кроссинговера в процессе мейоза. 5. Частота кроссинговера, определяемая по проценту кроссоверных особей, зависит от расстояния между генами. 6. На основании линейного расположения генов в хромосоме и частоты кроссинговера как показателя расстояния между генами можно построить карты хромосом. Сцепление генов приводит к одновременному наследованию блока признаков. Эти блоки могут сохраняться на протяжении нескольких поколений. Именно они способствуют передаче сложных признаков, например определяют внешнее сходство дедов и внуков, или обусловливают то, что все метисы колли или кокеров имеют совершенно определенную внешность независимо от внешнего вида второго родителя. Сцепление признаков может происходить в силу целого ряда причин, среди них и сцепление генов и плейотропия и действие определенных каналов морфогенеза. Сцепление признаков в значительной степени обусловливают тип собаки, препотентность производителя и делают возможным существование заводских линий. Кроссинговер (от англ. crossing–over – перекрёст) – это обмен гомологичными участками гомологичных хромосом (хроматид). Само явление перекрёста хромосом открыл Ф. Янссенс (1909, Бельгия), обнаруживший хиазмы в профазе I мейоза у саламандры. Однако теоретическая возможность кроссинговера была предсказана раньше. Во-первых, американский цитолог У. Сэттон (1903) предположил, что в одной хромосоме может находиться несколько генов. В этом случае должно наблюдаться сцепленное наследование признаков, т.е. несколько разных признаков могут наследоваться так, как будто они контролируются одним геном. Тогда совокупность генов в одной хромосоме образует группу сцепления. Во-вторых, в 1906 г. У. Бэтсон и Р. Пеннет обнаружили сцепленное наследование у душистого горошка. Они изучали совместное наследование: окраски цветков (пурпурная или красная) и формы пыльцевых зерен (удлиненная или округлая). При скрещивании дигетерозигот в их потомстве наблюдалось расщепление 11,1:0,9:0,9:3,1 вместо ожидаемого 9:3:3:1. Таким образом, два признака (окраска цветков и морфология пыльцы) оказываются сцепленными. Соответственно, сцеплены и гены, контролирующие эти признаки («эффект взаимного притяжения»). Однако это сцепление не абсолютно: гены, контролирующие сцепленные признаки, могут приобретать независимость, и тогда появляются новые комбинации признаков. Вскоре (1911) были получены данные по сцепленному наследованию и нарушению сцепления некоторых признаков у дрозофилы. Т. Морган (1910) и его ученик К. Бриджес (1916) доказали, что число групп сцепления соответствует гаплоидному числу хромосом. Опираясь на работы Янссенса и результаты гибридологического анализа, они связали явление перекрёста хромосом с феноменом нарушения сцепления генов. При перекрёсте хромосом происходит обмен между ними генетическим материалом (аллелями), и тогда происходит рекомбинация – появление новых сочетаний аллелей, например, AB + ab → Ab + aB. Таким образом, термин «кроссинговер» употребляется и в цитологическом, и в генетическом значении. В дальнейшем связь между перекрестом хромосом и появлением новых сочетаний признаков была подтверждена работами К. Дарлингтона, К. Штерна, Б. Мак-Клинток и др. Цитогенетика кроссинговера Механизм кроссинговера «разрыв–воссоединение» Согласно теории Янссенса–Дарлингтона, кроссинговер происходит в профазе мейоза. Гомологичные хромосомы с гаплотипами хроматид АВ и ab образуют биваленты. В одной из хроматид в первой хромосоме происходит разрыв на участке А–В, тогда в прилежащей хроматиде второй хромосомы происходит разрыв на участке a–b. Клетка стремится исправить повреждение с помощью ферментов репарации–рекомбинации и присоединить фрагменты хроматид. Однако при этом возможно присоединение крест–накрест (кроссинговер), и образуются рекомбинантные гаплотипы (хроматиды) Abи аВ. В анафазе первого деления мейоза происходит расхождение двухроматидных хромосом, а во втором делении – расхождение хроматид (однохроматидных хромосом). Хроматиды, которые не участвовали в кроссинговере, сохраняют исходные сочетания аллелей. Такие хроматиды (однохроматидные хромосомы) называются некроссоверными; с их участием разовьются некроссоверные гаметы, зиготы и особи. Рекомбинантные хроматиды, которые образовались в ходе кроссинговера, несут новые сочетания аллелей. Такие хроматиды (однохроматидные хромосомы) называются кроссоверными, с их участием разовьются кроссоверные гаметы, зиготы и особи. Таким образом, вследствие кроссинговера происходит рекомбинация – появление новых сочетаний (гаплотипов) наследственных задатков в хромосомах. 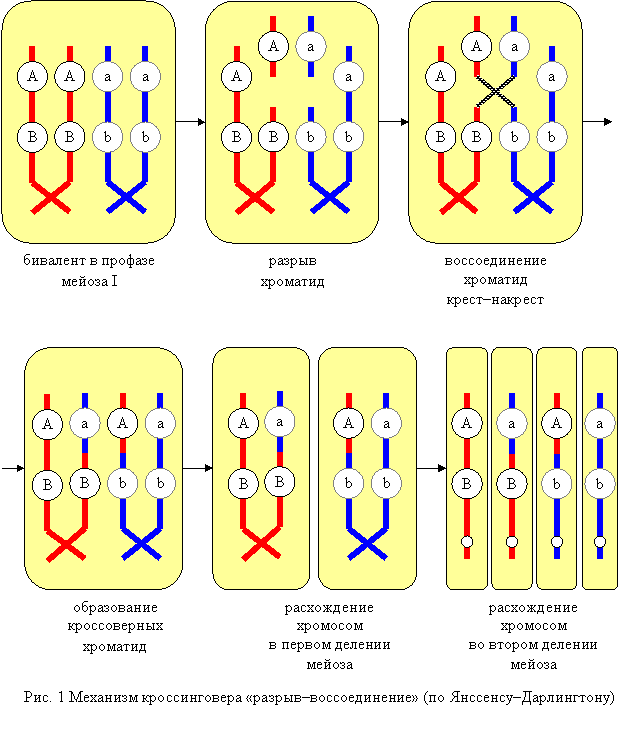 Примечание. Согласно другим теориям, кроссинговер связан с репликацией ДНК: или в пахитене мейоза, или в интерфазе (см. ниже). В частности, возможна смена матрицы в вилке репликации. Интерференция – это подавление кроссинговера на участках, непосредственно прилегающих к точке происшедшего обмена. Рассмотрим пример, описанный в одной из ранних работ Моргана. Он исследовал частоту кроссинговера между генами w (white – белые глаза), у (yellow – желтое тело) и m (miniature – маленькие крылья), локализованными в Х-хромосоме D. melanogaster. Расстояние между генами w и у в процентах кроссинговера составило 1,3, а между генами у и m – 32,6. Если два акта кроссинговера наблюдаются случайно, то ожидаемая частота двойного кроссинговера должна быть равна произведению частот кроссинговера между генами уи w и генами w и m. Другими словами, частота двойных кроссинговеров будет 0,43%. В действительности в опыте был обнаружен лишь один двойной кроссинговер на 2205 мух, т. е. 0,045%. Ученик Моргана Г. Меллер предложил определять интенсивность интерференции количественно, путем деления фактически наблюдаемой частоты двойного кроссинговера на теоретически ожидаемую (при отсутствии интерференции) частоту. Он назвал этот показатель коэффициентом коинциденции, т. е. совпадения. Меллер показал, что в Х-хромосоме дрозофилы интерференция особенно велика на небольших расстояниях; с увеличением интервала между генами интенсивность ее уменьшается и на расстоянии около 40 морганид и более коэффициент коинциденции достигает 1 (максимального своего значения). Двойной и множественный кроссинговер Т. Морган предположил, что кроссинговер между двумя генами может происходить не только в одной, но и в двух и даже большем числе точек. Четное число перекрестов между двумя генами, в конечном счете, не приводит к их перемещению из одной гомологичной хромосомы в другую, поэтому число кроссинговеров и, следовательно, расстояние между этими генами, определенное в эксперименте, снижаются. Обычно это относится к достаточно далеко расположенным друг от друга генам. Естественно, что вероятность двойного перекреста всегда меньше вероятности одинарного. В принципе она будет равна произведению вероятности двух единичных актов рекомбинации. Например, если одиночный перекрест будет происходить с частотой 0,2, то двойной – с частотой 0,2 × 0,2 = 0,04. В дальнейшем, наряду с двойным кроссинговером, было открыто и явление множественного кроссинговера: гомологичные хроматиды могут обмениваться участками в трех, четырех и более точках. Цитологическое (цитогенетическое) доказательство кроссинговера Прямые цитологические свидетельства обмена частей хромосом во время кроссинговера были получены в начале 30-х годов у дрозофилы и кукурузы. Рассмотрим опыт Штерна, проведенный на D. melanogaster. Обычно две гомологичные хромосомы морфологически неразличимы. Штерн исследовал Х-хромосомы, которые имели морфологические различия и, следовательно, были гомологичны не полностью. Однако гомология между этими хромосомами сохранялась на большей части их длины, что позволяло им нормально спариваться и сегрегировать в мейозе (то есть нормально распределяться по дочерним клеткам). Одна из Х-хромосом самки в результате транслокации, т. е. перемещения фрагмента Y-хромосомы, приобрела Г-образную форму. Вторая Х-хромосома была короче нормальной, так как часть ее была перенесена на IV хромосому. Были получены самки, гетерозиготные по указанным двум, морфологически различным, Х-хромосомам, а также гетерозиготные по двум генам, локализованным в Х-хромосоме: Bar (В) и carnation (cr). Ген Bar – это полудоминантный ген, влияющий на количество фасеток и, следовательно, форму глаза (мутанты с аллелем В имеют полосковидные глаза). Ген cr контролирует окраску глаз (аллель cr+ обусловливает нормальную окраску глаз, а аллель cr – окраску глаз цвета красной гвоздики). Г-образная Х-хромосома несла аллели дикого типа В+ и cr+, укороченная хромосома – мутантные аллели В и cr. Самки указанного генотипа скрещивались с самцами, имевшими морфологически нормальную Х-хромосому с аллелями cr и В+. В потомстве самок было два класса мух с некроссоверными хромосомами (crB / crB+ и cr+B+ / crB+) и два класса мух, фенотип которых соответствовал кроссоверам (crB+ / crB+ и cr+B / crB+). Цитологическое исследование показало, что у кроссоверных особей произошел обмен участками Х-хромосом, и, соответственно, изменилась их форма. Все четыре класса самок имели по одной нормальной, т. е. палочковидной, хромосоме, полученной от отца. Кроссоверные самки содержали в своем кариотипе преобразованные в результате кроссинговера Х-хромосомы – длинную палочковидную или двуплечую с короткими плечами. Эти опыты, так же как и одновременно полученные аналогичные результаты на кукурузе, подтвердили гипотезу Моргана и его сотрудников о том, что кроссинговер представляет собой обмен участками гомологичных хромосом и что гены действительно локализованы в хромосомах. 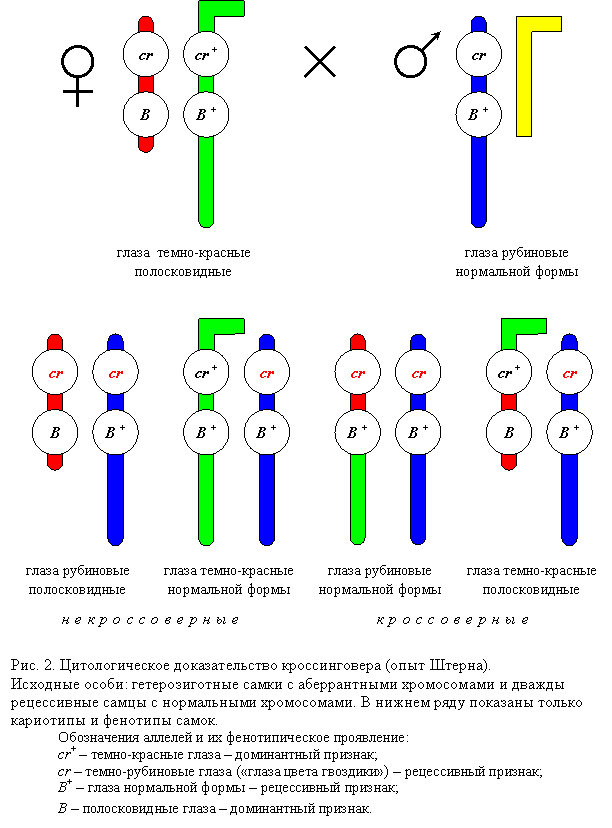 Соматический (митотический) кроссинговер В соматических клетках иногда происходят обмены между хроматидами гомологичных хромосом, в результате которых наблюдается комбинативная изменчивость, подобная той, которая регулярно генерируется мейозом. Нередко, особенно у дрозофилы и низших эукариот, гомологичные хромосомы синаптируют в митозе. Одна из аутосомно-рецессивных мутаций человека, в гомозиготном состоянии приводящая к тяжелому заболеванию, известному под названием синдром Блюма, сопровождается цитологической картиной, напоминающей синапсис гомологов и даже образование хиазм. Доказательство митотического кроссинговера было получено на дрозофиле при анализе изменчивости признаков, определяемых генами у (yellow – желтое тело) и sn (singed – опаленные щетинки), которые находятся в Х-хромосоме. Самка с генотипом ysn+ /y+sn гетерозиготна по генам у и sn, и поэтому в отсутствие митотического кроссинговера ее фенотип будет нормальным. Однако если кроссинговер произошел на стадии четырех хроматид между хроматидами разных гомологов (но не между сестринскими хроматидами), причем место обмена находится между геном sn и центромерой, то образуются клетки с генотипами y sn+ / y+ sn+ и y+sn/y+n. В этом случае на сером теле мухи с нормальными щетинками появятся близнецовые мозаичные пятна, одно из которых будет желтого цвета с нор мальными щетинками, а другое – серого цвета с опаленными щетинками. Для этого необходимо, чтобы после кроссинговера обе хромосомы (бывшие хроматиды каждого из гомологов) y+ sn отошли к одному полюсу клетки, а хромосомы y sn+ – к другому. Потомки дочерних клеток, размножившись на стадии куколки, и приведут к появлению мозаичных пятен. Таким образом, мозаичные пятна образуются тогда, когда рядом расположены две группы (точнее, два клона) клеток, фенотипически отличающиеся друг от друга и от клеток остальных тканей данной особи. |