доклады. Что изучает анатомия и физиология как наука, их значение Анатомия
 Скачать 0.81 Mb. Скачать 0.81 Mb.
|

|
Понятие нервных центров Нервный центр — центральный компонент рефлекторной дуги, где происходит переработка информации, вырабатывается программа действия, формируется эталон результата. Анатомическое понятие «нервный центр» — это совокупность нейронов, располагающихся в строго определенных отделах центральной нервной системы и осуществляющих один рефлекс. Например: центр коленного рефлекса — в передних рогах 2-4 поясничных сегментов спинного мозга; центр глотания — на уровне продолговатого мозга: 5, 7, 9 пары черепно-мозговых нервов. Физиологическое понятие «нервный центр» — это совокупность нейронов, расположенных на различных уровнях центральной нервной системы и регулирующих сложный рефлекторный процесс. Например: центр глотания входит в состав пищевого центра. Свойства нервных центров Одностороннее проведение возбуждения — возбуждение передается с афферентного на эфферентный нейрон. Причина: клапанное свойство синапса. Задержка проведения возбуждения: скорость проведения возбуждения в нервном центре на много ниже таковой по остальным компонентам рефлекторной дуги. Чем сложнее нервный центр, тем дольше проходит по нему нервный импульс. Причина: синаптическая задержка. Время проведения возбуждения через нервный центр — центральное время рефлекса. Суммация возбуждения — при действии одиночного подпорогового раздражителя ответной реакции нет. При действии нескольких подпороговых раздражителей ответная реакция есть. Рецептивное поле рефлекса — зона расположения рецепторов, возбуждение которых вызывает определенный рефлекторный акт. Имеется 2 вида суммации: временная и пространственная. Временная суммация — возникает ответная реакция при действии нескольких следующих друг за другом раздражителей. Механизм: суммируются возбуждающие постсинаптические потенциалы рецептивного поля одного рефлекса. Происходит суммация во времени потенциалов одних и тех же групп синапсов. Пространственная суммация — возникновение ответной реакции при одновременном действии нескольких подпороговых раздражителей. Механизм: суммация возбуждающего постсинаптического потенциала от разных рецептивных полей. Суммируются потенциалы разных групп синапсов. Центральное облегчение — объясняется особенностями строения нервного центра. Каждое афферентное волокно входя в нервный центр иннервирует определенное количество нервных клеток. Эти нейроны — нейронный пул. В каждом нервном центре много пулов. В каждом нейронном пуле — 2 зоны: центральная (здесь афферентное волокно над каждым нейроном образует достаточное для возбуждения количество синапсов), периферическая или краевая кайма (здесь количество синапсов недостаточно для возбуждения). При раздражении возбуждаются нейроны центральной зоны. Центральное облегчение: при одновременном раздражении 2-х афферентных нейронов ответная реакция может быть больше арифметической суммы раздражения каждого из них, т. к. импульсы от них отходят к одним и тем же нейронам периферической зоны. Окклюзия — при одновременном раздражении 2-х афферентных нейронов ответная реакция может быть меньше арифметической суммы раздражения каждого из них. Механизм: импульсы сходятся к одним и тем же нейронам центральной зоны. Возникновение окклюзии или центрального облегчения зависит от силы и частоты раздражения. При действии оптимального раздражителя, (максимального раздражителя (по силе и частоте) вызывающего максимальную ответную реакцию) — появляется центральное облегчение. При действии пессимального раздражителя (с силой и частотой вызывающих снижение ответной реакции) — возникает явление окклюзии. Посттетаническая потенция — усиление ответной реакции, наблюдается после серии нервных импульсов. Механизм: потенциация возбуждения в синапсах; Рефлекторное последействие — продолжение ответной реакции после прекращения действия раздражителя:
Трансформация возбуждения — несоответствие ответной реакции частоте наносимых раздражений. На афферентном нейроне происходит трансформация в сторону уменьшения из-за низкой лабильности синапса. На аксонах эфферентного нейрона, частота импульса больше частоты наносимых раздражений. Причина: внутри нервного центра образуются замкнутые нейронные цепи, в них циркулирует возбуждение и на выход из нервного центра импульсы подаются с большей частотой. Высокая утомляемость нервных центров — связана с высокой утомляемостью синапсов. Тонус нервного центра — умеренное возбуждение нейронов, которое регистрируется даже в состоянии относительного физиологического покоя. Причины: рефлекторное происхождение тонуса, гуморальное происхождение тонуса (действие метаболитов), влияние вышележащих отделов центральной нервной системы. Высокий уровень обменных процессов и, как следствие, высокая потребность в кислороде. Чем больше развиты нейроны, тем больше необходимо им кислорода. Нейроны спинного мозга проживут без кислорода 25-30 мин, нейроны ствола головного мозга — 15-20 мин, нейроны коры головного мозга — 5-6 мин. 24.Биотоки Биоэлектрические потенциалы (биотоки) — это электрические явления, наблюдаемые в живых клетках в покое и при физиологической деятельности. Возникновение в живых клетках электрических потенциалов и обусловленных ими биотоков связано с физико-химическими свойствами клеточных мембран и компонентов цитоплазмы (аминокислот, белков, ионов). Между наружной поверхностью клеточной мембраны и внутренним содержимым клетки существует всегда разность потенциалов, которая создается в силу различной концентрации ионов К+, Na+, Cl- внутри и вне клетки и различной проницаемости для них клеточной мембраны. Эта разность потенциалов называется «током покоя», или мембранным потенциалом, и составляет в среднем 60—90 мВ. При возбуждении живой клетки происходят изменения исходного мембранного потенциала за счет изменения проницаемости мембраны и перемещения ионов. В клетках возбудимых тканей (мышечной, нервной) эти процессы могут происходить в очень короткие интервалы времени (миллисекунды) и называются «током действия». Величина его может достигать 120 мВ. Для отведения биотоков от отдельных клеток организма попользуют специальные стеклянные и металлические микроэлектроды, имеющие неизолированные кончики (1—2 мк). Регистрация биоэлектрических потенциалов осуществляется с помощью катодных осциллографов и различных чернильнопишущих устройств с использованием усилителей биопотенциалов. См. также Электроэнцефалография, Электрокардиография, Электромиография, Электроретинография. Биоэлектрические явления (биоэлектрические потенциалы, биотоки) — электрические процессы, характерные для живых тканей. Биоэлектрические явления открыты Гальвани (A. L. Galvani) и Маттеуччи (С. Matteucci). Первые гипотезы о природе биоэлектрических явлений выдвинуты Дюбуа-Реймоном (Е. Du Bois-Reymond) и Германом (L. Hermann). В. Ю. Чаговец разработал теорию биоэлектрических явлений на основе современной физической химии электролитов. Подробное изучение биоэлектрических явлений стало возможным лишь с созданием соответствующей измерительной аппаратуры (катодные и шлейфные осциллографы в сочетании с высокочувствительными электронными усилителями). В настоящее время разработаны методы, позволяющие вводить электроды в глубь отдельных клеток живой ткани и регистрировать их электрическую активность. Такие исследования показывают, что между наружной и внутренней поверхностями клеточной мембраны существует постоянная разность потенциалов, причем наружная поверхность по отношению к внутренней имеет положительный заряд. Величина такой «трансмембранной» разности потенциалов составляет несколько десятков милливольт; она обозначается как «мембранный потенциал» или «потенциал покоя». Изменения потенциала покоя при различных функциональных состояниях клетки имеют названия: «потенциалы действия», «синаптические потенциалы», «генераторные потенциалы», «секреторные потенциалы» и т. д. Основой постоянной электрической поляризации поверхностной мембраны клетки является неравномерное распределение неорганических ионов (в первую очередь калия, натрия и хлора) между протоплазмой клетки и ее средой. В протоплазме клетки имеется значительный избыток ионов калия при сравнительно небольшом количестве ионов натрия и хлора (так называемая ионная асимметрия). В покоящемся состоянии клетки распределение ионов стационарно и поддерживается при неодинаковой проницаемости клеточной мембраны к различным ионам деятельностью активных протоплазматических механизмов, откачивающих определенные ионы из клетки или, наоборот, втягивающих их внутрь нее,— так называемый калиево-натриевый «насос», или «помпа». Подробно механизм деятельности этого «насоса» пока не выяснен, однако установлено, что он связан с источниками метаболической энергии клетки, особенно с системой расщепления макроэргич. фосфорных соединений. Ионы натрия и хлора оказывают влияние на потенциал покоя лишь при низких концентрациях ионов калия вне клетки. Наиболее точное определение величины разности потенциалов, существующей на клеточной поверхности, может быть проведено микроэлектродным методом, когда один из отводящих электродов (стеклянная микропипетка с диаметром кончика менее 0,5 мк) введен внутрь клетки, а вторым электродом служит окружающая клетку ткань. У млекопитающих, например, потенциал покоя нервной клетки составляет 60—80 мВ, мышечного поперечнополосатого волокна — 80—90 мВ, сердечного — 90—95 мВ. При возбуждении клетки изменяется ионная проницаемость ее поверхностной мембраны, вследствие чего появляются кратковременные перемещения ионов через мембрану. Эти ионные токи являются причиной изменений электрической поляризации мембраны клетки. Механизм возникновения наиболее распространенной формы активной электрической реакции — потенциала действия, связанного с распространяющейся волной возбуждения,— можно представить следующим образом. Необходимым условием возникновения распространяющегося возбуждения является снижение величины потенциала покоя (деполяризация) до определенной величины (истинный порог возбуждения клетки). Механизм возбуждения деполяризацией мембраны универсален; такое возбуждение возникает не только при электрическом, но и при любых других видах раздражения, в том числе при адекватных раздражениях рецепторных окончаний. Когда деполяризация достигает критического уровня (различного для разных типов клеток), стремительно развивается кратковременное повышение ионной проницаемости клеточной мембраны для таких ионов, которые в покоящемся состоянии с трудом проходят через мембрану. Природа этих изменений неизвестна; установлено, что в большинстве случаев ионом, который начинает особенно легко проходить через клеточную поверхность, является натрий. Положительные заряды ионов, движущихся внутрь клетки, не только полностью устраняют потенциал покоя, но даже на короткий момент так извращают трансмембранную разность потенциалов, что наружная поверхность мембраны становится отрицательной по отношению к внутренней ее стороне. В итоге этих процессов на поверхности клетки создается продольная разность потенциалов — возбужденный ее участок оказывается отрицательно заряженным по отношению к невозбужденному. Возникающие в связи с этим кольцевые электрические (ионные) токи между участком возбуждения и соседними невозбужденными участками («токи действия») являются причиной деполяризации невозбужденных участков до порогового уровня, что и обеспечивает распространение волны возбуждения по клетке. Амплитуда потенциала действия и его длительность у нервной клетки млекопитающих соответственно составляют 100—110 мВ и 1—2 мсек, у поперечнополосатого мышечного волокна — 110—120 мВ и 3—5 мсек. Однако потенциал действия у сердечного мышечного волокна оказывается чрезвычайно длительным. У каждой клетки амплитуда потенциала действия в нормальных условиях постоянна независимо от условий раздражения (правило «все или ничего»); однако если потенциал покоя по каким-либо причинам чрезвычайно снижен, то и амплитуда потенциала действия начинает уменьшаться либо генерация последнего оказывается совершенно невозможной («катодическая депрессия» или «инактивация»). После окончания основной «высоковольтной» части потенциала действия (пик) происходят дополнительные незначительные по амплитуде колебания электрической поляризации клеточной поверхности (так называемые следовые потенциалы). Выраженность их весьма варьирует у различных типов клеток. В большинстве случаев сразу после пика развивается следовая деполяризация, сменяющаяся следовой гиперполяризацией, которая, например, в соме нервной клетки достигает 100 мсек. Развитию процесса возбуждения соответствует и определенная теплопродукция, которая точно изучена на изолированном нерве и составляет в I стадии (начальное теплообразование) 7·10-8 кал на 1 г нерва на импульс, а во II стадии (отсроченное теплообразование) значительно превышает эту величину. В большинстве случаев развитию потенциала действия предшествуют промежуточные формы электрической активности. Одной из таких форм является локальный (местный) потенциал, возникающий при околопороговых раздражениях и отличающийся от потенциала действия градуальной зависимостью от силы раздражения (т. е. от величины вызывающей его деполяризации клеточной мембраны). В случае синаптической передачи возбуждения возникает особая форма локального потенциала — постсинаптический потенциал (ПСП), который также характеризуется градуальностью и способностью суммироваться с другими аналогичными потенциалами. ПСП является результатом специфической реакции постсинаптической мембраны клетки (т. е. той части ее мембраны, к которой прилегают синаптические окончания аксонов других клеток) на действие выделяемого синаптическими окончаниями медиатора. В зависимости от характера этого медиатора, а также, по-видимому, и от особенностей тех рецепторных группировок постсинаптической мембраны, которые с ним реагируют, ПСП могут выражаться изменениями электрической поляризации мембраны в различных направлениях. В одних случаях мембрана деполяризуется; если деполяризация достигает порогового значения, то генерируется обычный потенциал действия. Такие ПСП являются возбуждающими (ВПСП) и связаны с деятельностью особых возбуждающих синаптических окончаний. В других случаях электрическая поляризация мембраны увеличивается, а возникновение потенциала действия соответственно затрудняется; такие ПСП оказываются тормозящими (ТПСП) и лежат в основе синаптического торможения. При адекватном возбуждении нервных окончаний в рецепторах внешняя энергия трансформируется первоначально в градуальную деполяризацию их мембраны (генераторный потенциал), которая уже непосредственно приводит к развитию распространяющихся нервных импульсов, если раздражение достигает пороговой величины. Особенности генераторных потенциалов хорошо изучены на механорецепторах, простейших фоторецепторах и др. Секреторные процессы также связаны с возникновением на клеточной мембране железистой клетки градуальных секреторных потенциалов, которые могут иметь различные направление и длительность в зависимости от характера секреторной деятельности. Биоэлектрические потенциалы отдельных клеток могут суммироваться в электрические реакции целой ткани или органа. Так как ткань является проводником электричества (второго рода), то эти реакции могут быть зарегистрированы даже при расположении отводящих электродов на некотором расстоянии от органа (например, на коже). В нормальных условиях потенциал покоя клеток не обнаруживается в неповрежденной ткани, так как наружная поверхность каждой из них является изопотенциальной и имеет разность потенциалов только по отношению к внутреннему содержимому клетки. Однако если участок ткани повредить, разрушив тем или иным образом оболочки части клеток, то этот «альтерированный» участок во всех случаях окажется отрицательно заряженным по отношению к неповрежденным участкам той же ткани. Если достаточно большое количество клеток исследуемой ткани возбуждается одновременно и генерирует потенциалы действия или другие электрические реакции, то соответствующая электрическая реакция может быть зарегистрирована от целой ткани (нерва, мышцы, железы, участка мозга и т. д.). Как при повреждении, так и при возбуждении амплитуда колебаний будет значительно меньше, чем на мембране отдельной клетки, в связи с шунтированием внеклеточных токов межклеточной жидкостью. Во всех случаях электрод, соприкасающийся с возбужденным участком ткани, будет отрицательным по отношению к электроду, расположенному на невозбужденной части. Если импульс возбуждения поочередно проходит под одним и другим отводящими электродами, то будет зарегистрирован двухфазный ток действия; если же второй электрод расположить так, чтобы волна возбуждения не могла его достичь, то зарегистрированное колебание окажется однофазным. В неоднородной по клеточному составу ткани возникают более сложные типы электрических реакций, которые могут состоять из большого количества колебаний. Таким является, например, потенциал действия нервного ствола, содержащего волокна с различной скоростью проведения, если его отвести на некотором удалении от места раздражения, когда нервные импульсы в различных типах волокон уже успевают значительно разойтись во времени. От очень сложных по структуре образований, например от ЦНС, электрические колебания отводятся все время, даже при отсутствии специальных раздражений. Это связано с непрерывным поступлением импульсации от большого количества неконтролируемых источников, возможностью длительной циркуляции нервных импульсов по сложным цепям нейронов, а также генерацией ритмических импульсов нервными клетками под влиянием химических и других факторов. Электрическая активность соответствующих участков мозга проявляется ритмическими колебаниями очень небольшой амплитуды (несколько десятков мкв), отражающими общее функциональное состояние нейронов в соответствующей области мозга (хотя механизмы появления ритмических колебаний потенциала и изменения ритма при различных функциональных состояниях мозга пока не ясны). Биотоки, отведенные от поверхности коры больших полушарий, получили название электроэнцефалограммы; их регистрируют и изучают для физиологических и клинических исследований деятельности головного мозга (см. Электроэнцефалография). При раздражении различных сенсорных систем (рецепторы, афферентные нервы) в определенных областях мозга регистрируются характерные биоэлектрические ответы (так называемые вызванные потенциалы). Они имеют четкую локализацию на различных уровнях ЦНС и отражают некоторые характерные параметры возбудимых систем (латентный период). Путем регистраций вызванных потенциалов изучают пути передачи сенсорных импульсаций и их кортикальные проекции (локализацию). Широкое применение нашла регистрация суммарных электрических реакций сердечной мышцы (электрокардиография), скелетной мускулатуры (электромиография), сетчатой оболочки (электроретинография). 25.Пищеварение в тонком кишечнике.  Еда из желудка попадает в тонкий кишечник, если конкретнее - в двенадцатиперстную кишку. Двенадцатиперстная кишка – самый толстый отдел тонкого кишечника человека, ее длина около 30 см. Также к тонкому кишечнику относят тощую кишку (длина около 2,5м), подвздошную кишку (длина около 3 м). Внутренние стенки двенадцатиперстной кишки по своей сути состоят из множества мелких ворсинок. Под слоем слизи находятся небольшие железы, фермент которых способствует расщеплению белков. углеводов. Именно здесь жиры, белки. углеводы под действием пищеварительных соков, ферментов расщепляются таким образом, чтобы организм мог их легко усвоить. В двенадцатиперстную кишку, прежде всего, открывается протока поджелудочной железы, также желчный проток. Так что, на пищу здесь воздействуют:
Типы пищеварения в тонком отделе кишечника Контактное пищеварение: с помощью ферментов (мальтаза, сахараза) происходит расщепление до простых частиц, таких как аминокислоты и моносахариды. Происходит такое расщепление непосредственно в самом отделе тонкого кишечника. Но при этом остаются и мелкие частицы пищи, которые были расщеплены действием кишечного сока, желчи, но не достаточно, чтобы они могли усвоиться организмом. Такие частицы попадают в полость между ворсинками, которые плотным слоем покрывают слизистую в этом отделе. Здесь осуществляется пристеночное пищеварение. Концентрация ферментов здесь значительно выше. И поэтому, таким образом, процесс заметно убыстряется. Начальным предназначением ворсинок, кстати, было увеличение общей площади всасывающей поверхности. Длина двенадцатиперстной кишки достаточно мала. Перед тем как пища окажется в толстом кишечнике организму нужно успеть взять все питательные вещества из перерабатываемой пищи. Всасывание тонкого кишечника  Благодаря огромному количеству различных ворсинок, складок и отделов, а также особому строению выстилающих эпителиальных клеток, кишечник за час может всасывать до 3 литров потребляемой жидкости (как потребленной в чистом виде, так и с пищей). Все вещества, которые так попадают в кровь, транспортируются по вене в отдел печени. Это, безусловно, важно для организма, по той уже причине, что с пищей могут быть потреблены не только полезные вещества, но и различные токсины, яды – это связано, прежде всего, с экологией, а также с большим приемом лекарств, некачественной пищи и т.д. В отделах печени такая кровь обеззараживается и очищается. За 1 минуту печень способна переработать до 1,5 литров крови. В завершение через сфинктер остатки не переработанной пищи из подвздошной кишки попадают в толстый кишечник, а там уже происходит завершающий процесс пищеварения, а именно формирование каловых масс. Надо также заметить, что в толстом кишечнике пищеварение уже практически не происходит. Переваривается в основном только клетчатка и то также под действием ферментов, полученных в тонком кишечнике. Длина толстого кишечника – до 2 метров. В толстом кишечнике по факту происходит в основном лишь формирование кала и брожение. Вот почему так важно следить за здоровьем и нормальным функционированием тонкого кишечника, так как если возникают какие-то проблемы с двенадцатиперстной кишкой, то переработка потребленной пищи не будет завершена должным образом и, соответственно, организм недополучит целый ряд питательных веществ. Три пункта, воздействующие на усвоение пищи 1. Кишечный сок Вырабатывается непосредственно железами самого тонкого кишечника и дополняется своим действием общий процесс пищеварения этого отдела. По консистенции кишечный сок – это бесцветная мутноватая жидкость, с примесью слизи, а также эпителиальных клеток. Имеет щелочную реакцию. В состав входит более 20 важнейших пищеварительных ферментов (аминопептидаз, дипептидаз). 2. Поджелудочный (панкреатический) сок 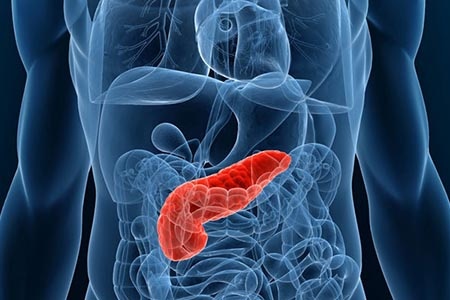 Поджелудочная железа – вторая по величине в организме человека. Вес может достигать 100г, а длина – 22 см. По сути поджелудочная железа делится на 2 отдельные железы:
Поджелудочный сок по своей сути – прозрачная бесцветная жидкость, имеющая рН 7,8 - 8,4. Выработка панкреатического сока начинается через 3 минуты после еды, а длится 6-14 часов. Больше всего поджелудочного сока выделяется при употреблении сильно жирной еды. Эндокринная железа синтезирует одновременно несколько гормонов, оказывающих важное действие на перерабатываемую пищу:
Ученые установили, что активность ферментов и их количественный состав в организме человека напрямую зависит от рациона человека. Чем больше он употребляет какой-то определенной еды – тем больше вырабатывается ферментов, необходимых именно для ее расщепления. 3. Желчь Самая большая железа в организме любого человека – печень. Именно она отвечает за синтез желчи, которая в дальнейшем накапливается желчным пузырем. Объем желчного пузыря сравнительно не большой – около 40 мл. Желчь в этом отделе организма человека содержится в очень концентрированном виде. Ее концентрация примерно в 5 раз выше, чем вырабатываемая изначально печеночная желчь. Просто все время в организм из нее всасываются минеральные соли и вода, а остается только концентрат, который имеет густую зеленоватую консистенцию с большим количеством пигментов. Поступать в отдел тонкого кишечника человека желчь начинает примерно через 10 минут после еды и вырабатывается, пока пища пребывает в желудке. Желчь не только оказывает воздействие на расщепление жиров и всасывание жирных кислот, но также еще повышает секрецию поджелудочного сока и улучшает перистальтику в каждом отделе кишечника. В отделы кишечника здорового человека в день выделяется до 1 литра желчи. Она состоит в основном из жиров, холестерина, слизи, мыла и лецитина. Как уже упоминалось ранее, проблемы с тонким кишечником способны привести к ужасным последствиям - организм будет недополучать полезные вещества, необходимые для нормальной жизнедеятельности организма. Вот почему так важно выявить любую проблему на ранней стадии, чтобы как можно быстрее начать ее лечение. Итак, возможные заболевания тонкого кишечника:
Причины возникновения заболеваний тонкого кишечника Иногда заболевания тонкого кишечника могут быть связанны с возрастными изменениями, наследственной предрасположенностью либо же врожденной патологией. Но есть и ряд провоцирующих факторов, которые по возможности следует исключить из жизни, чтобы предотвратить в дальнейшем проблемы со здоровьем:
Тошнота, рвота, диарея, слабость, боль в животе – наиболее ярко выраженные симптомы патологий, после обнаружения которых, следует безотлагательно обратиться к врачу. Чем быстрее будет диагностирована болезнь, а затем и начато лечение – тем выше вероятность в скором времени забыть о проблеме без каких-либо последствий для организма. 26.Мочевые органы. Образование мочи . |