Гиста 2022. Эпителиальная, ткани внутренней среды, мышечная и нервная ткани
 Скачать 5.1 Mb. Скачать 5.1 Mb.
|

|
Функции: каркас, обеспечивают упругость клетки, поддерживают упорядоченность расположения У человека множество ресничек имеют клетки эпителия бронхов. Они заставляют постоянно двигаться вверх слой слизи с частицами пыли и остатками отмерших клеток. С помощью ресничек клеток яйцевода яйцеклетки продвигаются по нему. Жгутики отличаются от ресничек лишь длиной. Так, сперматозоиды млекопитающих ( рис. 75 ) имеют по одному жгутику длиной до 100 мкм. Обычно реснички короче жгутиков более чем в 10 раз. Тысячи ресничек одной клетки движутся координированно, образуя на поверхности плазмалеммы бегущие волны ( рис. 29 ). Каждая ресничка работает подобно хлысту: удар вперед, при котором ресничка полностью выпрямляется и передает в окружающую жидкость максимальное усилие, проталкивая ее, а затем, изгибаясь, чтобы уменьшить сопротивление среды, она возвращается в исходное положение 16. Микроворсинки. Строение, функции. Примеры локализации Микроворсинки — это цилиндрические пальцеообразные выросты цитоплазмы, покрытые плазмолеммой. В частности, они присутствуют на апикальной поверхности эпителиальных клеток тонкой кишки, т. е. обращены в просвет этой кишки. Форма и положение микроворсинок поддерживаются благодаря наличию в них, как и в других областях цитоплазмы, цитоскелета. В данном случае он представлен микрофиламентами— нитями из белка актина, которые расположены вдоль оси микроворсинки. Микроворсинки сливаются в непрерывную и узкую оксифильную каемку на апикальной поверхности клеток. Данная каемка называется щеточной или всасывающей, а соответствующие клетки — каемчатыми. Около 30 параллельно идущих микрофиламентов образуют стержень микроворсинки. (+)-концы двух переплетенных нитей F-актина микрофиламентов направлены к вершине микроворсинки. Микрофиламенты сшивают актин-связывающие белки фимбрин и виллин. Микрофиламенты присоединены к внутренней поверхности плазматической мембраны при помощи миозина I. В основании микроворсинок актиновые нити (-)-концами заякорены в терминальную сеть – примембранное сплетение микрофиламентов сшитых между собой фодрином. Каждый энтероцит (осуществляющая всасывание эпителиальная клетка тонкой кишки) содержит более 1000 микроворсинок, которые увеличивают площадь апикальной поверхности клетки в 20 раз. Пример: микроворсинки тонкой кишки, осуществляющие всасывание. 17. Классификация межклеточных контактов. Примеры локализации различных контактов Некоторые адгезивные белки плазмолеммы клеток участвуют в образовании долговременных межклеточных соединений (контактов). Межклеточные контакты (рис. 2.12) по своим функциональным свойствам подразделяются на четыре группы: 1) контакты простого типа — простые межклеточные соединения и интердигитации (пальцевидные соединения); необходимы для механического сцепления клеток друг с другом. Это просто сближение плазмолемм соседних клеток на расстояние 15–20 нм без образования специальных структур. При простом соединении плазмолеммы взаимодействуют друг с другом с помощью специфических адгезивных гликопротеинов — кадгеринов, интегринов и др. Плазмолемма двух клеток при интердигитации, сопровождая друг друга, инвагинирует в цитоплазму вначале одной, а затем — соседней клетки. 2) контакты сцепляющего типа — десмосомы и адгезивные пояски; так же необходимы для механического сцепления клеток друг с другом. I. В области десмосомы плазмолеммы обеих клеток с внутренней стороны утолщены — за счет белков десмоплакинов, образующих дополнительный слой. От этого слоя в цитоплазму клетки отходит пучок промежуточных филаментов, участвующих в образовании цитоскелета. II. В области десмосомы плазмолеммы обеих клеток с внутренней стороны могут быть заполнены утолщенным гликокаликсом, который пронизан сцепляющими белками — десмоглеинами. Последние образуют фибриллоподобные структуры и дисковидное утолщение посередине. Если клетка лежит на базальной мембране, то связь между ними (клеткой и мембраной) осуществляется с помощью полудесмосом. Это значит, что со стороны клетки присутствуют все элементы десмосомы (и внутри-, и вне-клеточные), включая дисковидную пластинку. Но теперь эта пластинка прикреплена непосредственно к базальной мембране. Адгезивный поясок по структуре похож на десмосомный, но имеет ряд отличительных черт. Прежде всего это касается формы контакта: представляет собой ленту, которая опоясывает клетку. Иная и природа белков: I. вместо десмоплакинов используется винкулин (утолщения плазмолемм со стороны цитоплазмы); II. вместо промежуточных филаментов — тонкие филаменты, образованные во всех клетках белком актином (нити, отходящие в цитоплазму); III. вместо десмоглеинов — другие сцепляющие белки (в пространстве между плазмолеммами). 3) контакты запирающего типа — плотное соединение (запирающая зона, или zona occludens); полное разграничение сред, лежащих по разные стороны клеточного пласта. Здесь плазмолеммы прилегают друг к другу вплотную, сцепляясь с помощью специальных белков. Места такого плотного прилегания образуют на контактирующих поверхностях подобие ячеистой сети. 4) контакты коммуникационного типа — щелевидные соединения (нексусы, или gap-junctions) и синапсы. Позволяют клеткам обмениваться веществами (нексусы) или сигналами (синапсы). Щелевидное соединение (нексус) имеет форму круга диаметром 0,5–3 мкм. Плазмолеммы контактирующих клеток сближены здесь на расстояние 2 нм и пронизаны многочисленными полыми трубочками — каналами (с просветом в 2 нм), которые связывают цитоплазму клеток. Каждая трубочка состоит из двух половин — коннексонов. Коннексон пронизывает мембрану лишь одной клетки и выступает в межклеточную щель на 1 – 1,5 нм, где стыкуется со вторым коннексоном. Через образуемые коннексонами каналы могут диффундировать неорганические ионы и большинство низкомолекулярных органических соединений — сахара, аминокислоты, промежуточные продукты их метаболизма. Это означает, что между контактирующими клетками существует электрическая и метаболическая связь. Ионы Са2+ меняют конфигурацию коннексонов — так, что просвет каналов закрывается. Синапсы служат для передачи сигнала от одних возбудимых клеток к другим. В этом отношении их функция отчасти сходна с функцией нексусов. Но структура синапса совсем иная. В синапсе различают: – пресинаптическую мембрану (ПреМ), принадлежащую одной клетке, – синаптическую щель – постсинаптическую мембрану (ПоМ) - часть плазмолеммы другой клетки. Обычно сигнал передается химическим веществом — медиатором (4): последний диффундирует от ПреМ и воздействует на специфические рецепторы в ПоМ. 18. Десмосома и полудесмосома. Строение, значение, примеры локализации В области десмосомы плазмолеммы обеих клеток с внутренней стороны утолщены — за счет белков десмоплакинов, образующих дополнительный слой. От этого слоя в цитоплазму клетки отходит пучок промежуточных филаментов, участвующих в образовании цитоскелета. II. В области десмосомы плазмолеммы обеих клеток с внутренней стороны могут быть заполнены утолщенным гликокаликсом, который пронизан сцепляющими белками — десмоглеинами. Последние образуют фибриллоподобные структуры и дисковидное утолщение посередине. Если клетка лежит на базальной мембране, то связь между ними (клеткой и мембраной) осуществляется с помощью полудесмосом. Это значит, что со стороны клетки присутствуют все элементы десмосомы (и внутри-, и вне-клеточные), включая дисковидную пластинку. Но теперь эта пластинка прикреплена непосредственно к базальной мембране. Десмосома представляет собой небольшое округлое образование, содержащее специфические структурные элементы. Плазматические мембраны клеток разделены промежутком 20-30 нм, в котором находятся внемембранные части Са-связывающих белков десмоглеина и десмоколлина. К внутренней поверхности плазматической мембрнаы прилегает цитоплазматическая пластинка с вплетенными в нее промежуточными филаментами. В состав этой пластинки входят десмоплакины, плакоглобин и часть молекулы десмоглеина. Два компонента: - цитоплазматическая пластинка (с вплетенными в нее промежуточными филаментами) - десмоглея (внеклеточный мембрнанный материал) Полудесмосома. Обеспечивает прикрепление клетки к базальной мембране. Как и десмосома, содержит цитоплазматическую пластинку с вплетенными в нее промежуточными филаментами. Например, прикрепление кератиноцитов базального слоя эпидермиса. участвующих в образовании цитоскелета. В области десмосомы плазмолеммы обеих клеток с внутренней стороны могут быть заполнены утолщенным гликокаликсом, который пронизан сцепляющими белками — десмоглеинами. Последние образуют фибриллоподобные структуры и дисковидное утолщение посередине. Если клетка лежит на базальной мембране, то связь между ними (клеткой и мембраной) осуществляется с помощью полудесмосом. Это значит, что со стороны клетки присутствуют все элементы десмосомы (и внутри-, и вне-клеточные), включая дисковидную пластинку. Но теперь эта пластинка прикреплена непосредственно к базальной мембране. 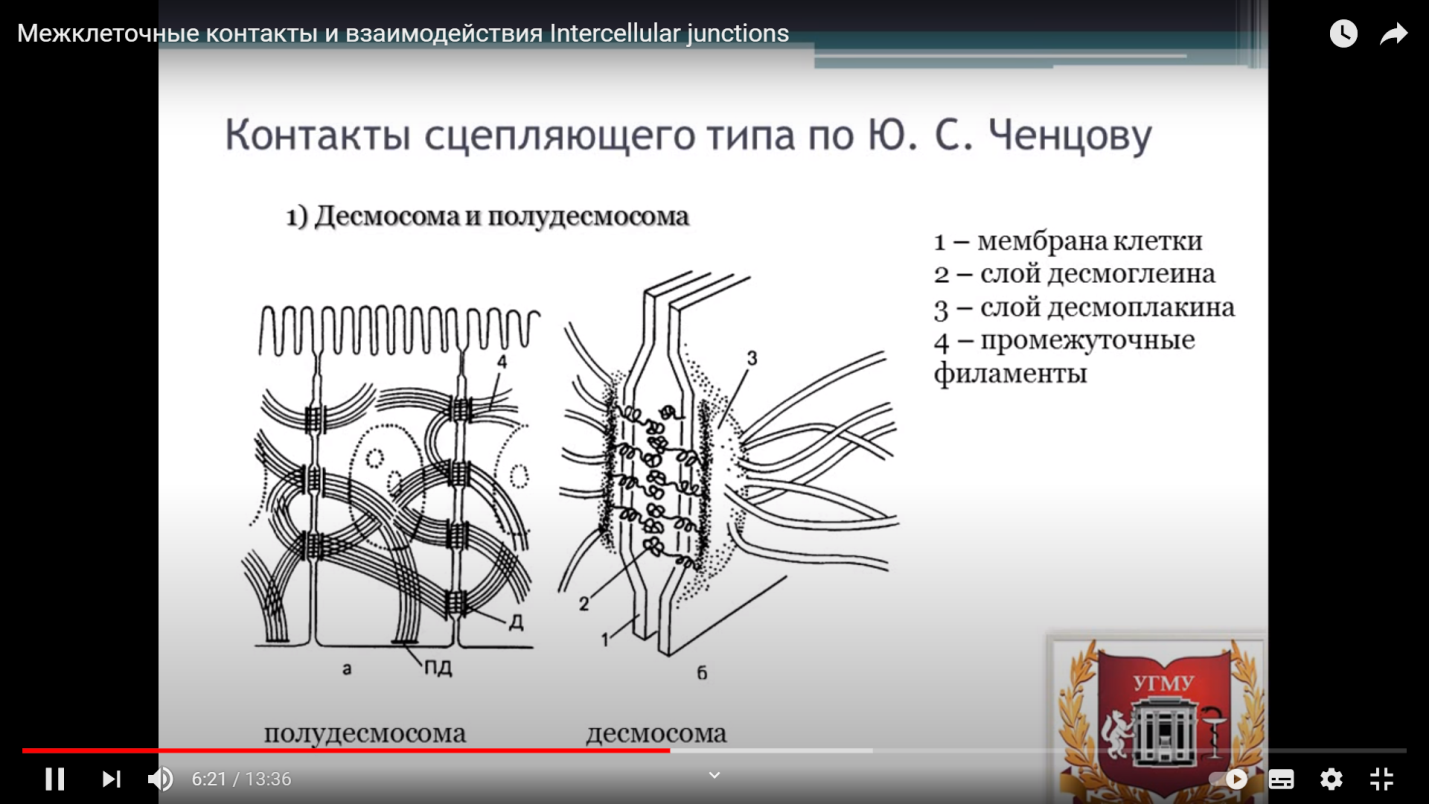 19. Щелевой контакт. Структура, функция, примеры локализации Щелевидное соединение (нексус) имеет форму круга диаметром 0,5–3 мкм. Плазмолеммы контактирующих клеток сближены здесь на расстояние 2 нм и пронизаны многочисленными полыми трубочками — каналами (с просветом в 2 нм), которые связывают цитоплазму клеток. Каждая трубочка состоит из двух половин — коннексонов. Коннексон – трансмембранный белок цилиндрической конфигурации, состоящий из 6 субъединиц коннексина. Коннексон пронизывает мембрану лишь одной клетки и выступает в межклеточную щель на 1 – 1,5 нм, где стыкуется со вторым коннексоном. Через образуемые коннексонами каналы могут диффундировать неорганические ионы и большинство низкомолекулярных органических соединений — сахара, аминокислоты, промежуточные продукты их метаболизма. Это означает, что между контактирующими клетками существует электрическая и метаболическая связь. Ионы Са2+ меняют конфигурацию коннексонов — так, что просвет каналов закрывается. Функция: проведение низкомолекулярных соединений, регулирующих рост и развитие клеток. Обеспечение распространения возбуждения – переход ионов между мышечными клетками миокарда и между ГМК. 20. Плотные контакты. Строение, локализация, функции Места такого плотного прилегания образуют на контактирующих поверхностях подобие ячеистой сети. Плотный контакт (или замыкающий) формирует в клеточных слоях регулируемый барьер проницаемости, разделяющий разные по химическому составу среды (например, внутреннюю и внешнюю). Плотные контакты выглядят как ленты, состоящие из цепочек округлых зон (бляшек) размером 10 нм и с расстоянием между центрами соседних бляшек примерно 18 нм. Здесь плазмолеммы прилегают друг к другу вплотную, сцепляясь с помощью специальных белков. Белки, составляющие многомолекулярный комплекс плотного контакта, можно разделить на три группы: интегральные, белки бляшки контакта, цитоплазматические белки. Примеры локализации: каемчатый эпителий кишки, эндотелий капилляров, альвеолоциты, эпителиальные клетки почечных канальцев. 21. Пиноцитоз: характеристика, примеры Пиноцитоз — захват и поглощение клеткой растворов веществ (в составе капельки), с образованием небольших пузырьков. Подразделяется на макропиноцитоз (диаметр эндосом 0,2-0,3 мкм) и микропиноцитоз (диаметр эндосом 70-100 нм). Пиноцитоз рассматривается как неспецифический способ полощения внеклеточных жидкостей и содержащихся в ней веществ, когда некоторая область клеточной мембраны впячивается, образует ямку и далее пузырек, содержащий межклеточную жидкость. Пузырьки, которые при этом образуются, часто бывают очень мелкими. В таком случае говорят о микропиноцитозе и пузырьки называют микропиноцитозными. Пиноцитоз характерен для очень многих клеток, как животных, так и растительных. Примеры: захват эпителиальными клетками кишечника и почечных канальцев, эндотелием сосудов. 22. Аксо-вазальные синапсы. Локализация, строение, функции Аксо-вазальные синапсы - это окончания аксонов нейросекреторных нейронов на капиллярах. Локализация – в гипоталамо-гипофизарной системе - в стенках гемокапилляров нейрогемальных органов - в стенках желудочков и каналов мозга -в оболочках мозга Функция: секреция вазопрессина и окситоцина в капилляры задней доли гипофиза, секреция рилизинг-гормонов и капилляры срединного возвышения, выведение нейрогормонов в кровь или лимфу Образуются нейронами гипоталамуса. Строение: • Концевые ветвления эфферентного нейрона образуют подошвообразные или булавовидные расширения • В окружении терминалей отсутствуют леммоциты • В аксоплазме терминалей находятся синаптические пузырьки с медиатором или нейрогормоном (секреторное нейроны) • Терминали проникает через базальную мембрану и контактирует с цитолеммой иннервируемой клетки или образуют сплетения под базальной мембраной • Между аксолеммой и плазмолеммой имеется щель (100 нм), в которую выбрасывается медиатор • На цитолемме иннервируемой клетки расположены рецепторы к медиаторам 23. Холинергический синапс, строение Холинергические синапсы — синапсы, в которых передача возбуждения осуществляется посредством ацетилхолина. При связывании АХ с никотиновым холинрецептором (н-холинрецептор) в составе последнего открывается ионный канал, через пору которого проходят ионы натрия и калия, приводя к деполяризации постсинаптической мембраны (постсинаптический потенциал). Холинергический синапс включает 5 белковых субъединиц (α, α, β, γ, δ), окружающих ионный (натриевый) канал и проходящих через всю толщу липидной мембраны. Две молекулы ацетилхолина взаимодействуют с двумя α-субъединицами, что приводит к открыванию ионного канала и деполяризации постсинаптической мембраны. Холинорецепторы разной локализации обладают неодинаковой чувствительностью к фармакологическим веществам. На этом основано выделение так называемых мускариночувствительных холинорецепторов — м-холинорецепторы (мускарин — алкалоид из ряда ядовитых грибов, например мухоморов) и никотиночувствительных холинорецепторов — н-холинорецепторы (никотин — алкалоид из листьев табака). В синапсах с н-холинорецепторами реализуется ионотропный механизм: – рецепторы связаны с катионными каналами, которые открываются под действием ацетилхолина; – при этом внутрь постсинаптической клетки начинают поступать (по градиенту своей концентрации) ионы Na+, – поэтому развивается деполяризация, а значит, возбуждение постсинаптической мембраны. м-Холинергические синапсы являются метаботропными. В этом случае связывание медиатора с рецептором меняет внутри постсинаптической клетки (через мембранные и внутриклеточные посредники) активность регуляторных ферментов — чаще всего протеинкиназ. Последние же фосфорилируют определенные белки, влияя таким образом уже на их активность (повышая ее или понижая). Таким образом, один и тот же нейромедиатор (как в данном случае ацетилхолин) иногда действует в одних синапсах по ионотропному, а в других синапсах — по метаботропному механизму. 24. Нервно-мышечный синапс. Строение, функционирование, нейромедиатор Нервно-мышечный синапс (моторная бляшка)-двигательное окончание аксона мотонейрона на волокнах поперечнополосатых соматических мышц-состоит из концевого ветвления аксона, образующего пресинаптическую часть, специализированного участка на мышечном волокне, соответствующего постсинаптической части, и разделяющей их синаптической щели. Пресинаптическая часть содержит синаптические пузырьки с нейромедиатором (в каждом пузырьке – квант нейромедиатора), элементы цитоскелета и митохондрии. В пресинаптическую мембрану встроены потенциалозависимые Са-каналы. Синаптическая щель – промежуток между мембранами шириной 20 – 35 нм. В синаптическую щель из синаптических пузырьков выделяются молекулы нейромедиатора и путем диффузии достигают постсинаптической мембраны. В синаптической щели находятся ферменты, расщепляющие молекулы нейромедиатора (АХэстераза, гидролизирующая АХ), а в пресинаптическую мембрану вмонтированы переносчики, осуществляющие перенос нейромедиаторов-аминокислот и биогенных аминов (глутамата, аспартата, норадреналина) в пресинаптическую терминаль. Постсинаптическая область – много крупных митохондрий с хорошо развитыми кристами и большое количество рибосом. На постсинаптической мембране располагаются рецепторы к нейромедиатору ацетилхолину. 25. Строение сперматозоида В эякуляте человека содержится 3*108 сперматозоидов. В женских половых путях они сохраняют способность к оплодотворению максимум двое суток. Примерно 200 из них достигает воронки маточной трубы. В сперматозоиде различают две части: головку и хвост, или жгутик. Головка сперматозоида человека имеет форму эллипсоида, сжатого с боков, с одной из сторон имеется небольшая ямка. Акросома образуется в ходе сперматогенеза как продукт комплекса Гольджи.Акросома расположена в головке сперматозоида, кпереди от ядра и сразу под плазматической мембраной. Спереди мембрана акросомы (наружная) соприкасается с клеточной мембраной сперматозоида, а сзади (внутренняя мембрана) – с ядерной мембраной. Акросома содержит ферменты, необходимые для проникновения сперматозоида в яйцеклетку: акрозин, пенетразу, гиалуронидазу, коллагеназу, кислую фосфатазу и др. Главный компонент головки — клеточное ядро с гаплоидным набором конденсированных хромосом. Содержит в основании ядерные белки уже не прежние гистоны, а другие основные белки (гистоны и протамины), более богатые аргинином и цистеином. Плазмолемма головки. Вся головка сперматозоида, как и его хвост, окружена плазматической мембраной. В области головки эта мембрана содержит специальные белки: одни из них заряжены отрицательно и способствуют (на малых расстояниях) направленному движению сперматозоида к яйцеклетке; другие белки участвуют в связывании с яйцеклеткой. От головки среднюю часть отделяет небольшое сужение — «шейка». Короткая шейка содержит две центриоли — проксимальную и дистальную, образованные микротрубочками |