ответы экзамен. Эволюция пищеварительной системы и особенности питания у представителей разных классов позвоночных животных
 Скачать 7.56 Mb. Скачать 7.56 Mb.
|

|
высвободившийся в синаптическую щель медиатор диффундирует к постсинаптической мембране и вступает во взаимодействие с ее хеморецепторами, функционально связанными с воротным механизмом хемовозбудимых каналов; в результате такого взаимодействия медиатора с хеморецепторами постсинаптической мембраны срабатывают (как правило, открытием) определенные хемовозбудимые каналы и временно изменяется проницаемость постсинаптической мембраны (в ней отсутствуют потенциалзависимые каналы) для определенных ионов. Изменение проницаемости постсинаптической мембраны для определенных ионов, вызванное действием медиатора, носит дискретный характер и будет продолжаться до тех пор, пока медиатор не расщепится соответствующим ферментом, всегда присутствующим в синаптической щели химических синапсов. Постсинаптические мембраны различных химических синапсов центральной нервной системы могут характеризоваться различной избирательной проницаемостью своих хемовозбудимых каналов для определенных ионов. Так, если постсинаптическая мембрана химического синапса содержит хемовозбудимые каналы, проницаемые преимущественно для ионов Na+или Са2+, то открытие этих каналов под действием присоединившегося к хеморецептору медиатора будет сопровождаться усиленным током положительных зарядов в постсинаптический полюс, и как следствие, деполяризацией постсинаптической мембраны (возникает возбуждающий постсинаптический потенциал, ВПСП). Деполяризация постсинаптической мембраны химического синапса, связывающего какие-то нейроны, может послужить предпосылкой последующей генерации нервного импульса в области аксонного холмика иннервируемого нейрона. В связи с отмеченным химические синапсы, в которых медиатор вызывает деполяризацию постсинаптической мембраны (возникновение возбуждающего постсинаптического потенциала), являются возбуждающими с функциональной точки зрения. В случае, если постсинаптическая мембрана химического синапса содержит хемовозбудимые каналы, преимущественно проницаемые для ионов К+ или Cl-, то открытие таких каналов, возникающее в результате присоединения медиатора к хеморецептору, будет сопровождаться либо выходящим калиевым током, либо входящим хлорным, которые гиперполяризуют постсинаптическую мембрану. Гиперполяризация постсинаптической мембраны, в свою очередь, может снижать возбудимость определенного участка мембраны иннервируемого нейрона, либо ослаблять суммарную деполяризацию его мембраны, вызванную возбуждением, поступающим через другие синапсы возбуждающего типа, расположенные на нем. Таким образом, гиперполяризация постсинаптической мембраны снижает возбудимость иннервируемого нервного элемента (такой ответ постсинаптической мембраны получил название тормозного постсинаптического потенциала, ТПСП), а химические синапсы, в которых взаимодействие медиатора с хеморецепторами постсинаптической мембраны сопровождается ее гиперполяризацией, являются тормозными с функциональной точки зрения. Таким образом, синапсы с химическим способом передачи информации обеспечивают проведение информации всегда в одном направлении с некоторой синаптической задержкой и могут быть как возбуждающими,так итормознымис функциональной точки зрения. Синапсы электрического типа характеризуются очень узкой синаптической щелью (2-4 нм) и наличием белковых мостиков (образованы белком коннексином) между пре- и постсинаптической мембранами. Через такие белковые мостики способны легко диффундировать от пресинаптического полюса к постсинаптическому многие низкомолекулярные вещества (ионы, АТФ, глюкоза, аминокислоты и некоторые другие). Узкая синаптическая щель и наличие белковых мостиков делает возможным непосредственную передачу возбуждения (без участия химических посредников) от пресинаптической мембраны к постсинаптической мембране: ионные токи, возникающие при возбуждении пресинаптической мембраны, достигают, значительно не ослабевая при этом, постсинаптической мембраны электрического синапса, вызывая возникновение в ней всегда только деполяризации (возбуждающего постсинаптического потенциала). Постсинаптическая мембрана электрического синапса не содержит хемовозбудимых ионных каналов, но характеризуется наличием, как и все возбудимые структуры, потенциалзависимых каналов. Следовательно, электрические синапсы проводят информацию от пресинаптической мембраны на постсинаптическую без участия медиаторов и без синаптической задержки, в функциональном плане могут быть только возбуждающими. 34.Ультраструктурная организация нервной клетки. Нервные клетки (нейроны) состоят из тела (сома или перикарион) и отростков двух типов – дендритов (как правило, короткие, ветвящиеся, немиелинизированные) и аксона (нейрит, нервное волокно; одиночный, длинный отросток, может быть миелинизированным или немиелинизированным). Дендриты выполняют в нейроне воспринимающую функцию. В случае вставочных и эфферентных нейронов дендриты являются местом локализации большого количества .В случае чувствительных нейронов дендриты либо непосредственно образуют рецепторы, воспринимающие действие раздражителей, либо принимают участие в их образовании. Таким образом, дендриты чувствительных нейронов выступают в качестве собственно рецепторного аппарата нервной системы, воспринимая действие факторов внешней или внутренней среды организма. Тело нейрона (сома или перикарион) выполняет следующие функции: осуществление обменных процессов, необходимых для поддержания жизнедеятельности всей клетки (трофическая функция) восприятие информации, поступающей от других нейронов (на мембране тела, подобно таковой дендритов, также имеются сотни - тысячи синапсов, образуемых другими нейронами) интегративная функция(объединение той информации, которая поступает к дендритам и телу от других нейронов, и анализ этой информации). Аксон нейрона (нервное волокно или нейрит) специализируется на проведение информации в виде нервного импульса от тела клетки к другим структурам. В морфологическом плане нейроны классифицируют на: 1 2. Биполярные : 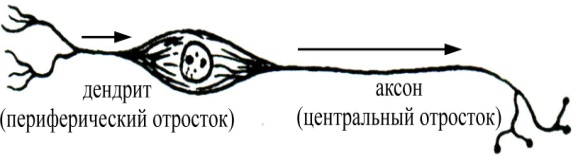 а) типичные биполярные - От тела клетки отходят два отростка, которые в морфологическом плане соответствуют аксону (т.е. являются, как правило, миелинизированными нервными волокнами, способными к генерации и проведению нервного импульса). В функциональном плане один отросток соответствует дендриту (проводит информацию к телу клетки), а другой – аксону (проводит информацию от тела нейрона к другим нейронам). Такие нейроны всегда являются чувствительными (обонятельные нейроны, нейроны вестибулярного и кортиевого ганглиев, биполярные нейроны сетчатки глазного яблока). а) типичные биполярные - От тела клетки отходят два отростка, которые в морфологическом плане соответствуют аксону (т.е. являются, как правило, миелинизированными нервными волокнами, способными к генерации и проведению нервного импульса). В функциональном плане один отросток соответствует дендриту (проводит информацию к телу клетки), а другой – аксону (проводит информацию от тела нейрона к другим нейронам). Такие нейроны всегда являются чувствительными (обонятельные нейроны, нейроны вестибулярного и кортиевого ганглиев, биполярные нейроны сетчатки глазного яблока).б 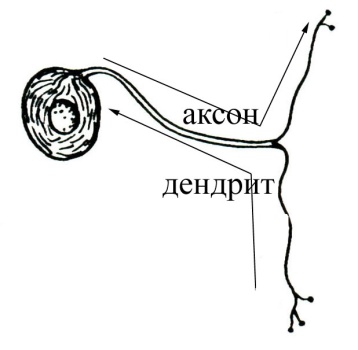 ) псевдоуниполярные - Развиваются в процессе эмбриогенеза из типичных биполярных нейронов путем схождения двух отростков в одну точку. Оба отростка имеют сходное морфологическое строение, но один отросток, функционально соответствующий дендриту, проводит информацию к телу клетки, а другой, функционально соответствующий аксону, – от тела клетки к другим нейронам. Такие нейроны всегда являются чувствительными(нейроны спинальных ганглиев, полулунного, коленчатого, каменистого и яремного ганглиев черепно-мозговых нервов). ) псевдоуниполярные - Развиваются в процессе эмбриогенеза из типичных биполярных нейронов путем схождения двух отростков в одну точку. Оба отростка имеют сходное морфологическое строение, но один отросток, функционально соответствующий дендриту, проводит информацию к телу клетки, а другой, функционально соответствующий аксону, – от тела клетки к другим нейронам. Такие нейроны всегда являются чувствительными(нейроны спинальных ганглиев, полулунного, коленчатого, каменистого и яремного ганглиев черепно-мозговых нервов).3 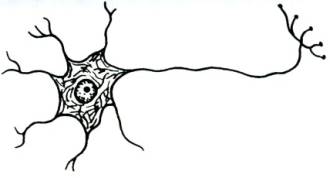 . Мультиполярные - Количество отростков – три и более. Такие нейроны, как правило, в функциональном плане являются вставочными и эфферентными, хотя могут быть и чувствительными (ганглиозные клетки сетчатки глазного яблока). . Мультиполярные - Количество отростков – три и более. Такие нейроны, как правило, в функциональном плане являются вставочными и эфферентными, хотя могут быть и чувствительными (ганглиозные клетки сетчатки глазного яблока).В функциональном плане нейроны можно классифицировать по: 1. локализации в составе рефлекторной дуги чувствительные (афферентные), находятся за пределами центральной нервной системы, но, как правило, вблизи неё, в чувствительных узлах спинномозговых или черепно-мозговых нервов, или в редких случаях на периферии там же, где и рецепторы. Дендриты следуют на периферию, где непосредственно могут образовывать рецепторы, либо принимают участие в их образовании, вступая в синаптический контакт со специальными ненервными рецепторными клетками (вторичночувствующие рецепторы). Аксоны чувствительных нейронов следуют в центральную нервную систему в составе чувствительных ветвей спинно- или черепномозговых нервов. вставочные (промежуточные, кондукторные), тела и отростки этих нейронов находятся в пределах центральной нервной системы; представляют собой вставочное звено на пути нервного импульса от чувствительных к эфферентным нейронам (т.е. передают информацию в пределах цнс). эфферентные, передают информацию из центральной нервной системы к рабочим органам – скелетным мышцам или внутренним органам, сосудам, гладкой мускулатуре и железам кожи. Тела соматических нейронов (мотонейронов) находятся в пределах центральной нервной системы, а их аксоны (эфферентные или двигательные нервные волокна) следуют к рабочим органам (скелетные мышцы). 2. Согласно принципу Дейла один и тот же нейрон во всех пресинаптических окончаниях своего аксона выделяет один и тот же медиатор. В связи с этим нейроны можно классифицировать по характеру выделяемого медиатора: холинергические адренергические дофаминергические серотонинергические нейросекреторные клетки (обнаруживаются в гипоталамусе, синтезируют и секретируют в кровь рилизинг-факторы, вазопрессин и окситоцин) и т.д. 3. электрофизиологическим свойствам возбуждающие (оказывают возбуждающее влияние на другие нейроны или структуры рабочего органа, с которыми вступают в синаптический контакт) тормозные (клетки Реншоу спинного мозга, клетки Пуркинье коры мозжечка, амакриновые нейроны сетчатки и другие), оказывают тормозное влияние на те структуры, с которыми вступают в синаптический контакт. Анатомия растений, Цитология и Ботаника 35.Формы полового процесса у грибов. Подразделение грибов на отделы. Половое размножение. Половым называется размножение, при котором имеет место половой процесс, завершающийся слиянием ядер специализированных клеток (гамет, гаметангиев) либо соматических клеток. У грибов отмечаются следующие формы полового процесса: Гаметогамия (планогамия). Это половой процесс, при котором сливаются две гаметы (одна или обе подвижные, одинаковые или разной величины). Подвижные гаметы называют планогаметами. Гаметогамия подразделяется на изогамию, при которой сливаются (копулируют) обе гаметы, подвижные и морфологически неразличимые, и гетерогамию (анизогамию), при которой сливаются (копулируют) подвижные, отличающиеся по размерам и часто по степени подвижности гаметы. Оогамия, при которой образуются половые структуры – оогоний и антеридий. В оогонии образуется неподвижная крупная яйцеклетка, а в антеридии формируются мелкие подвижные сперматозоиды, которые в итоге проникают в яйцеклетку и оплодотворяют ее. В результате образуется зигота (ооспора). Гаметангиогамия – половой процесс, при котором сливается содержимое двух специализированных половых структур – гаметангиев, не дифференцированных на гаметы. Гаметангии обычно многоядерные, и в результате их слияния наряду со слиянием цитоплазмы происходит множественное слияние ядер. У зигомицетов сливаются гаметангии, в большинстве случаев морфологически хорошо отличимые от гиф мицелия, на котором они образуются. В результате слияния формируется зигоспора, которая после периода покоя прорастает в спорангий. Соматогамия – процесс, при котором половые структуры не образуются, а сливаются обычные соматические или вегетативные клетки гиф мицелия. При этом сливаются две одноклеточные особи. Такая разновидность соматогамии называется хологамией. У зигомицетов гаметангиогамия называется зигогамией. Половой процесс происходит следующим образом: на разнополых (гетероталличных) или однополых (гомоталличных) мицелиях образуются особые боковые веточки (зигофоры) – слегка вздувшиеся на концах фрагменты гиф. Затем в каждом зигофоре с помощью поперечной стенки отделяется многоядерный (или одноядерный) гаметангий. Стенка между обоими гаметангиями разрушается, их протопласты сливаются (плазмогамия), ядра перемешиваются. Образовавшаяся в результате единая клетка окружается многослойной стенкой и созревает в покоящуюся спору, называемую зигоспорой или зигоспорангием. После длительного периода созревания (до нескольких месяцев) и покоя она прорастает. В это время часть ядер располагается попарно, а другие дегенерируют. Несколько пар ядер (иногда – одна) сливаются, происходит кариогамия, образуются диплоидные ядра. По истечении периода покоя зигоспора прорастает, при этом все диплоидные ядра, кроме одного, погибают (дегенерируют). Выжившее диплоидное ядро мейотически делится, из 4 образовавшихся гаплоидных ядер одно прорастает и в итоге формируется спорангиеносец со спорангием, в котором все спорангиоспоры однополые. Сперматизация. Некоторые грибы образуют мелкие одноядерные структуры наподобие конидий на конидиеносце, микроконидии, способные к копуляции, в общем называемые спермациями. У многих аскомикот вместо антеридия его функцию выполняют вегетативные клетки гиф, микроконидии, конидии или спермации, аскоспоры, почкующиеся клетки. Сумчатым грибам свойственна партеногамия – самооплодотворение внутри аскогона, а также апомиксис. Царство грибов делится на два отдела: 1. Оомикота. 2. Эумикота (настоящие грибы). Классы разграничиваются на основании типа органов размножения и ряда признаков строения вегетативного тела грибов. К отделу оомикота принадлежат только два класса (оомицеты и гифохитриомицеты). Они отличаются числом жгутиков и составом клеточных оболочек. Подавляющее большинство грибов (96%) относится к отделу Эумикота, в котором различают пять классов (хитридиомицеты, зигомицеты, аскомицеты, базидиомицеты, дейтеромицеты). Так называемые макромицеты – грибы с плодовыми телами курпных размеров, хорошо различимыми без микроскопа – это представители двух классов грибов – базидиомицетов и аскомицетов. 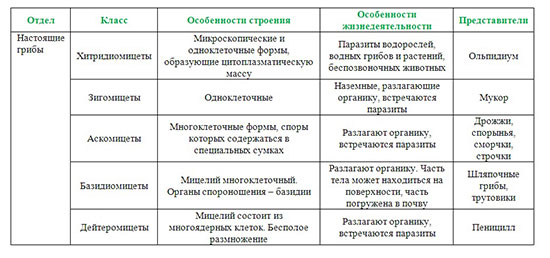 36.Растительные ткани, их классификация и функции. 36.Растительные ткани, их классификация и функции. Ткани – это устойчивые, генетически детерминированные комплексы клеток, сходных по происхождению, строению, местоположению и выполняемым функциям. Термин «ткани» был введен в ботанику Н. Грю (1671). Учение о тканях получило название гистология. К истокам гистологии растений следует отнести работу английского естествоиспытателя Р. Гука «Микрография» (1669), в которой впервые было дано описание срезов стебля бузины, укропа, тростника и других растений. Ткани делят на простые и сложные. Простыми называют ткани, состоящие из клеток более или менее одинаковых по форме и функциям. Сложные ткани состоят из клеток, разных по форме и функциям, но тесно взаимосвязанных в своих жизненных отправлениях. Пример первых — столбчатая хлоренхима, губчатая хлоренхима, колленхима, вторых — ксилема, флоэма. Ткани делятся на образовательные (меристема) и постоянные: Образовательными называются специализированные ткани, клетки которых сохраняют длительную способность к делению, обеспечивая рост растения и отдельных его органов. С учетом положения в теле растения их делят на апикальные (или верхушечные, находятся на апексах корня и побега), интеркалярные (или вставочные, свойственны побегу — стеблю и листьям, находятся в междоузлиях и черешках) и боковые (или латеральные, представлены главным образом в осевых органах — в корне и стебле голосеменных и двудольных покрытосемянных). Постоянными называют ткани, клетки которых утратили способность к делению (полностью или сохраняют её потенциально) и специализируются на выполнении других функций: защитной, запасающей, механической, проводящей и т. д. С учетом происхождения, преобладающей функции и положения в теле растения постоянные ткани, в свою очередь, делят на покровные, проводящие и основные, начало которым при первичном росте дают соответственно протодерма, прокамбий и основная меристема. Наряду с анатомо-физиологической существует и онтогенетическая классификация тканей, основанная на их происхождении и времени появления в процессе морфогенеза органа. По этой классификации ткани делят на первичные и вторичные. Первичные меристемы ведут свое начало от первой клетки нового организма — зиготы, которым свойственна способность к делению. Они первыми формируются при заложении нового организма и обеспечивают его первичный рост. Это — верхушечные и вставочные меристемы. Те постоянные ткани, клетки которых дифференцируются из производных клеток первичной меристемы, называют первичными. К ним относят ткани: первичные покровные, первично проводящие и основные. Вторичными называют меристемы, которые формируются в вегетативных органах позднее первичных и обеспечивают их вторичный рост. Это боковые меристемы — камбий и феллоген (пробковый камбий). Постоянные ткани, начало которым дали производные клетки вторичной меристемы, называют вторичными. К ним относятся вторичную покровную ткань, вторичные проводящие ткани. Классификация растительных тканей основана на единстве выполняемых функций, происхождении, сходстве строения и расположении клеток в органах растения. По этим критериям ткани делят на несколько групп: меристематические или образовательные, покровные, основные, механические, проводящие, выделительные.
|