ответы экзамен. Эволюция пищеварительной системы и особенности питания у представителей разных классов позвоночных животных
 Скачать 7.56 Mb. Скачать 7.56 Mb.
|

|
Генетика Закон Н.И. Вавилова о гомологической изменчивости. Николай Иванович Вавилов имел научные интересы в широких сферах, но более всего он известен как ботаник-селекционер и генетик. Исследуя параллелизм в наследственной изменчивости, в 1920 году он открыл и обосновал закон гомологических рядов, который звучит так: «Виды и роды, генетически близкие, характеризуются сходными рядами наследственной изменчивости с такой правильностью, что, зная ряд форм в пределах одного вида, можно предвидеть нахождение параллельных форм у других видов и родов. Чем ближе генетически расположены в общей системе роды и виды, тем полнее сходство в рядах их изменчивости. Целые семейства растений в общем характеризуются определенным циклом изменчивости, проходящей через все роды и виды, составляющие семейства». Для наглядной иллюстрации действия закона удобно взять растения семейства мятликовых. Например, зерновка черной окраски имеется у пшеницы, ячменя, ржи, кукурузы и ряда других представителей семейства. А вот удлиненная форма зерновки наблюдается у всех видов семейства, изученных на сегодняшний день. С помощью этого закона Н. И. Вавилов обнаружил ряд форм ржи, еще не известных в то время: с остистыми и безостными колосьями, с черной, белой, фиолетовой и красной зерновкой, с зерном мучнистым и стекловидным и проч. Исследователь опирался на наличие данных признаков у пшеницы. Он выявил базовые закономерности в биологии, сходные с теми строгими закономерностями, которые Менделеев обнаружил для химических элементов. Закон гомологических рядов наследственной изменчивости можно применять как для растений, так и для животных. Например: 1. Короткопалость встречается и у человека, и у птиц, собак, крупного рогатого скота. 2. Альбинизм бывает у представителей разных групп млекопитающих, а также у птиц и прочих животных. 3. Перья могут отсутствовать у птиц, чешуя у рыб, волосистый покров у млекопитающих, и всё это производные кератина. Закон, открытый Вавиловым, очень важен в селекции. Он дает возможность предугадать появление форм, доселе не обнаруженных у конкретного вида, однако имеющихся у близкородственных видов. Данную форму можно «вдруг» обнаружить в дикой природе, но можно и получить с помощью искусственного мутагенеза. Уровни упаковки генетического материала. Структурная организация хроматина. Хроматин имеет определенную пространственную организацию, характеризующуюся его компктизацией. Необходимость компактизации определяется большой длиной молекул ДНК ядра. Компактизация молекул ДНК, приводящая к возникновению, укорочению и утолщению хроматиновых фибрилл достигается путем взаимодействия ДНК с гистонами и белками ядерного матрикса. На этой основе формируются 3 универсальных, прогрессивно усложняющихся уровня пространственной организации хроматина: нуклеосмный, нуклеомерный и хромомерный. Нуклеосомный уровень организации хроматина характеризуется образованием нуклеосом. Нуклеосома – это комплекс участка молекулы ДНК с белками-гистонами. Белковой основой нуклеосомы является коровая частица, или сердцевина, представляющая собой гистоновый гетерооктамер, т.е. содержащая 8 молекул разных гистонов. Коровая частица взаимодействует с участком молекулы ДНК величиной 145-146 пар нуклеотидов, который обвивает поверхность сердцевины почти 2 раза. Такой комплекс участка ДНК с коровой частицей называют коровой нуклеосомой. С коровой нуклеосомой связывается одна молекула гистона Н1 (глобулярным доменом – с сердцевиной, концевыми доменами – с ДНК), в результате чего формируется полная нуклеосома, включающая 9 гистоновых молекул. На данном уровне длина нуклеосомной нити приблизительно в 7 раз меньше, чем длина соответствущей молекулы ДНК в неспирализованном состоянии. Нуклеосомный уровень организации не препятствует использованию нуклеосомной ДНК в качестве матрицы для синтеза ДНК и РНК. Нуклеомерный уровень организации хроматина формируется на основе нуклеосомного путем образования нуклеомер. Переход хроматина с нуклеосомного уровня на нуклеомерный осуществляется с помощью гистона Н1 в составе полной нуклеосомы. С помощью своих концевых доменов молекула гистона Н1 способна связываться с молекулами Н1 соседних нуклеосом. Такой комплекс и называют нуклеомерой. Образование нуклеомер приводит к формированию нуклеомерной нити хроматина толщиной 25-30 нм и компактизированной в 40 раз. На данном уровне ДНК теряет способность быть матрицей в процессах синтеза ДНК и РНК. Хромомерный уровень организации хроматина формируется взаимодействием специфических участков молекулы ДНК хромосом с белками ядерного матрикса с образованием хромомер. На этом уровне организации хроматиновые нити имеют толщину около 300 нм и компактизированы в тысячи раз. Функции ядерного матрикса. Главной функцией ядерного аппарата является обеспечение и регуляция потока генетической информации, заложенной в ДНК хроматина – генетическом материале. Это выражается в ее хранении, удвоении, исправлении и первых этапах реализации. Биохимическую основу потока информации составляют матричные процессы. Основные положения хромосомной теории наследственности. Суммируя данные о независимом и сцепленном наследовании признаков, о наследовании, сцепленных с полом, о связи нарушения поведения хромосом и отклонения от закономерностей наследования признаков у дрозофилы и других организмов, Морган и его сотрудники сформулировали основные положения хромосомной теории наследственности: - Гены расположены в хромосоме линейно. - Гены, расположенные в одной хромосоме, образуют группу сцепления, вследствие чего они наследуются вместе. - Между гомологичными хромосомами может происходить кроссинговер, благодаря чему образуются новые сочетания генов. - Сила сцепления обратно пропорциональна расстоянию между генами. Хромосомная теория наследственности (Х. т. н.), теория, согласно которой хромосомы, заключённые в ядре клетки, являются носителями генов и представляют собой материальную основу наследственности, т.е. преемственность свойств организмов в ряду поколений определяется преемственностью их хромосом. Х. т. н. возникла в начале 20 в. на основе клеточной теории и использования для изучения наследственных свойств организмов гибридологического анализа. В 1902 У. Сеттон в США, обративший внимание на параллелизм в поведении хромосом и менделевских т. н. «наследственных факторов», и Т. Бовери в Германии выдвинули хромосомную гипотезу наследственности, согласно которой менделевские наследственные факторы (название впоследствии генами) локализованы в хромосомах. Первые подтверждения этой гипотезы были получены при изучении генетического механизма определения пола у животных, когда было выяснено, что в основе этого механизма лежит распределение половых хромосом среди потомков. Дальнейшее обоснование Х. т. н. принадлежит американскому генетику Т. Х. Моргану, который заметил, что передача некоторых генов (например, гена, обусловливающего белоглазие у самок дрозофилы при скрещивании с красноглазыми самцами) связана с передачей половой Х-хромосомы, т. е. что наследуются признаки, сцепленные с полом (у человека известно несколько десятков таких признаков, в том числе некоторые наследственные дефекты — дальтонизм, гемофилия и др.). Доказательство Х. т. н. было получено в 1913 американским генетиком К. Бриджесом, открывшим нерасхождение хромосом в процессе мейоза у самок дрозофилы и отметившим, что нарушение в распределении половых хромосом сопровождается изменениями в наследовании признаков, сцепленных с полом. С развитием Х. т. н. было установлено, что гены, расположенные в одной хромосоме, составляют одну группу сцепления и должны наследоваться совместно; число групп сцепления равно числу пар хромосом, постоянному для каждого вида организмов; признаки, зависящие от сцепленных генов, также наследуются совместно. Вследствие этого закон независимого комбинирования признаков должен иметь ограниченное применение; независимо должны наследоваться признаки, гены которых расположены в разных (негомологичных) хромосомах. Явление неполного сцепления генов (когда наряду с родительскими сочетаниями признаков в потомстве от скрещиваний обнаруживаются и новые, рекомбинантные, их сочетания) было подробно исследовано Морганом и его сотрудниками (А. Г. Стёртевантом и др.) и послужило обоснованием линейного расположения генов в хромосомах. Морган предположил, что сцепленные гены гомологичных хромосом, Морган предположил, что сцепленные гены гомологичных хромосом, находящиеся у родителей в сочетаниях 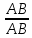 и и 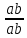 , в мейозе у гетерозиготной формы → , в мейозе у гетерозиготной формы → 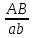 могут меняться местами, в результате чего наряду с гаметами АВ и ab образуются гаметы Ab и аВ. Подобные перекомбинации происходят благодаря разрывам гомологичных хромосом на участке между генами могут меняться местами, в результате чего наряду с гаметами АВ и ab образуются гаметы Ab и аВ. Подобные перекомбинации происходят благодаря разрывам гомологичных хромосом на участке между генами 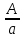 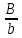 и последующему соединению разорванных концов в новом сочетании: и последующему соединению разорванных концов в новом сочетании: Реальность этого процесса, названного перекрестом хромосом, или кроссинговером, была доказана в 1933 нем, учёным К. Штерномв опытах с дрозофилой и американскими учёными Х. Крейтономи Б. Мак-Клинток — с кукурузой. Чем дальше друг от друга расположены сцепленные гены, тем больше вероятность кроссинговера между ними. Зависимость частоты кроссинговера от расстояний между сцепленными генами была использована для построения генетических карт хромосом. Согласно представлениям школы Моргана, гены являются дискретными и далее неделимыми носителями наследственной информации. Однако открытие в 1925 советскими учёными Г. А. Надсоном и Г. С. Филипповым, а в 1927 американским учёным Г. Мёллером влияния рентгеновских лучей на возникновение наследственных изменений (мутаций) у дрозофилы, а также применение рентгеновских лучей для ускорения мутационного процесса у дрозофилы позволили советским учёным А. С. Серебровскому, Н. П. Дубинину и др. сформулировать в 1928—30 представления о делимости гена на более мелкие единицы, расположенные в линейной последовательности и способные к мутационным изменениям. Использование рентгеновских лучей для стимулирования хромосомных перестроек позволило Н. П. Дубинину и Б. Н. Сидорову обнаружить в 1934 эффект положения гена, т. е. зависимость проявления гена от места расположения его на хромосоме. Возникло представление о единстве дискретности и непрерывности в строении хромосомы. Х. т. н., объясняя закономерности наследования признаков у животных и растительных организмов, играет важную роль в сельском хозяйстве, науке и практике. Она вооружает селекционеров методами выведения пород животных и сортов растений с заданными свойствами. Так, явление сцепленного с полом наследования ряда признаков у с.-х. животных позволило до изобретения методов искусственного регулирования пола у тутового шелкопряда выбраковывать коконы менее продуктивного пола, до разработки способа разделения цыплят по полу исследованием клоаки — отбраковывать петушков и т.п. Важнейшее значение для повышения урожайности многих с.-х. культур имеет использование полиплоидии. На знании закономерностей хромосомных перестроек основывается изучение наследственных заболеваний человека. Теория эволюции Значение теории Ч. Дарвина для фундаментальной науки. Формы изменчивости по Ч. Дарвину. Появление эволюционной теории Ч. Дарвина имело огромное значение для биологии. Развитие органического мира нашло в ней свое естественное материалистическое объяснение. Тем самым был нанесен непоправимый удар по церковным догмам о сотворении мира и его неизменности. Именно за эту диалектико-материалистическую сущность теория Дарвина была очень высоко оценена классиками марксизма-ленинизма. Теория Дарвина внесла в биологию исторический метод. Все биологические науки в теории Дарвина получили совершенно новую, истинно научную основу. Огромные новые перспективы открылись и перед селекцией. Вскрытые Дарвином факторы породо- и сортообразования позволили более широко и умело использовать их, ускоряя и совершенствуя селекционный процесс. Возникла специальная наука генетика, значительно расширившая знания о наследственности и изменчивости. Основываясь на эволюционном значении сложных взаимоотношений между организмами в природе, человек оказался вооруженным эффективными способами воздействия на природу. Наконец, именно благодаря теории Дарвина человек смог заглянуть в свое далекое прошлое. Основы антропологии – науки о происхождении человека — были заложены Ч. Дарвином. Если всеми прогрессивными учеными мира теория Дарвина была встречена с восторгом как долгожданная и необходимая основа для дальнейших исследований, то совершенно иначе отнеслись к этой материалистической теории церковь и реакционные ученые-идеалисты. Немало было попыток опровергнуть и исказить дарвинизм. Но все эти попытки наталкивались на твердый и решительный отпор со стороны последователей и защитников теории Дарвина. В России особенно много в этом отношении было сделано горячим пропагандистом Дарвина — К. А. Тимирязевым. Он взял на себя нелегкий труд сделать эту научную теорию достоянием широких масс общественности. Ему принадлежит блестящее по тонкости, научной строгости и предельной доступности изложение этой теории в книге «Чарльз Дарвин и его учение». Вместе с тем теория Дарвина стала основой и его собственных научных исследований в области физиологии растений. То же надо сказать и о других русских последователях Дарвина, принявших его теорию за основу своих исследований. К ним относятся эмбриологи А. О. Ковалевский и И. И. Мечников, палеонтолог В. О. Ковалевский и многие другие. Так эволюционная теория Чарльза Дарвина, созданная свыше ста лет назад, стала материалистической основой наших современных представлений о природе, о путях и закономерностях ее развития, о способах управления биологическими процессами. Ч. Дарвин установил, между особями одного вида всегда существуют как хорошо заметные различия (спорты), так и малозаметные. Ч. Дарвин считал, что именно мелкие различия между особями являются материалом для отбора. Эти различия Ч. Дарвин назвал изменчивостью и выделил несколько форм изменчивости: определенную, неопределенную, комбинативную и коррелятивную. Ненаследственная изменчивость является определенной, поскольку, изменяя условия развития организмов, можно предвидеть направление изменчивости. В то же время, она является и групповой, поскольку вся группа особей, подвергаемая одинаковому изменению условия развития, изменяется в одном направлении. [В XX веке такую изменчивость долгое время называли модификационной.] Например, если группу поросят одной породы выращивать в хороших условиях, то через полгода все они будут характеризоваться сходными чертами: большая масса (около 200 кг), удлиненное тело, укороченные конечности, слабо развитый шерстный покров, спокойное поведение, хороший аппетит. Если же группу поросят той же породы выращивать в плохих условиях, то взрослые особи также будут сходны между собой: низкий вес (около 50 кг), укороченное тело, удлиненные конечности, сильно развитый шерстный покров, злобный нрав, плохой аппетит. Наследственная изменчивость прямо противоположна ненаследственной. Эта изменчивость неопределенная. Например, мы не можем заранее предсказать: когда и в каком стаде появится овца с резко укороченными конечностями. Наследственная изменчивость является индивидуальной: изменение признака наблюдается лишь у одной особи из многих. [В XX веке эту форму изменчивости долгое время называли мутационной.] Ч. Дарвин противопоставлял наследственную изменчивость ненаследственной и считал, что к отбору приводит только наследственная изменчивость: «Ненаследственное изменение для нас несущественно». Коррелятивная изменчивость. Пример с изменением целого комплекса признаков у поросят показывает, что при изменении условий развития изменяется не один признак, а целый комплекс признаков. Одновременное изменение нескольких признаков Ч. Дарвин назвал соотносительной, или коррелятивной изменчивостью. Наследственная изменчивость также может быть коррелятивной: изменение одного признака влечет за собой изменение других признаков. Например, все белые голубоглазые коты – глухие, у бесшерстных собак определенной породы – недоразвиты зубы. [В настоящее время установлено, что наследственная коррелятивная изменчивость может быть обусловлена множественным действием генов, а также сцеплением генов – истинным сцеплением или квазисцеплением.] Комбинативная изменчивость. Во времена Дарвина (еще до опытов Менделя) было известно, что разные наследственные признаки могут образовывать различные комбинации (например, разнообразные сочетания окраски и длины шерсти у животных). Независимую изменчивость признаков Ч. Дарвин назвал комбинативной изменчивостью. [В настоящее время установлено, что коррелятивная изменчивость часто сочетается с комбинативной, т.е. совместная изменчивость двух и более признаков часто носит комбинативно-коррелятивный характер.] Отсутствие генетической теории в XIX веке привело Ч. Дарвина к некоторым ошибочным представлениям о наследственности. Например, Ч. Дарвин считал, что степень проявления признаков у потомков равна среднему арифметическому значению этого признака у родителей. Эта посылка («кошмар Дженкина») была серьезным препятствием для дальнейшего развития дарвинизма. Кроме того, Ч. Дарвин допускал возможность наследования благоприобретенных признаков, поддерживая теорию пангенезиса. Согласно этой теории, в крови существуют особые частицы – геммулы (буквально – «зачатки», «почки»), которые переносят информацию от всех частей тела к половым железам. В результате информация об онтогенезе особи может передаваться через кровь к половым клеткам. Заметим, что геммулы Дарвина сходны по своим свойствам с флюидами Ламарка. Ламаркизм Ч. Дарвина проявлялся и в частичном признании закона упражнения и неупражнения. Например, редукцию органов зрения у троглобионтов Ч. Дарвин объяснял ненужностью этого признака в темноте. +В 1900 г. происходит переоткрытие законов И.Г. Менделя, формирование представлений о дискретном характере наследственности и изменчивости. Тем самым был устранен «кошмар Дженкина». В 1901 г. Г. де Фриз создает мутационную теорию. К 1908 г. формируются основные представления о генетической структуре популяций (закон Харди-Вайнберга). Учение о наследственности до сих пор не завершено. Однако современная генетика утверждает, что любой признак в той или иной степени наследуется. Отрицать эволюционную роль ненаследственных изменений (модификаций) нельзя. «Все ненаследственное в своем возникновении связано с наследственным. Модификация организма всегда определяется его наследственной структурой» (Шмальгаузен). |