ответы экзамен. Эволюция пищеварительной системы и особенности питания у представителей разных классов позвоночных животных
 Скачать 7.56 Mb. Скачать 7.56 Mb.
|

|
76. Фотосинтетический цикл Хэтча-Слэка (С4-путь фотосинтеза). Первичный и конечный продукты фотосинтеза.  С4-фотосинтез (альтернативный путь фотосинтеза), или цикл Хэтча — Слэка, (Ю. С. Карпилов, 1960,1969; М. Д. Хэтч и С. Р. Слэк, 1966) - путь связывания углерода, характерный для высших растений, первым продуктом которого является четырёхуглеродная щавелевоуксусная кислота, а не трёхуглеродная 3-фосфоглицериновая кислота, как у большинства растений с обычным С3-фотосинтезом. В настоящее время С4 фотосинтез обнаружен у 943 видов растений, которые относятся к 18 семействам. Эти растения отличаются и анатомически: паренхима сосудисто- волокнистых пучков у них содержит хлоропласты, чего не имеют С3-растения. Во флоре территории СНГ найдено 130 видов таких растений, из них 60 видов относятся к сем. Маревых, 34 вида – к сем. Злаковых. В числе их есть С4-фотосинтез (альтернативный путь фотосинтеза), или цикл Хэтча — Слэка, (Ю. С. Карпилов, 1960,1969; М. Д. Хэтч и С. Р. Слэк, 1966) - путь связывания углерода, характерный для высших растений, первым продуктом которого является четырёхуглеродная щавелевоуксусная кислота, а не трёхуглеродная 3-фосфоглицериновая кислота, как у большинства растений с обычным С3-фотосинтезом. В настоящее время С4 фотосинтез обнаружен у 943 видов растений, которые относятся к 18 семействам. Эти растения отличаются и анатомически: паренхима сосудисто- волокнистых пучков у них содержит хлоропласты, чего не имеют С3-растения. Во флоре территории СНГ найдено 130 видов таких растений, из них 60 видов относятся к сем. Маревых, 34 вида – к сем. Злаковых. В числе их есть 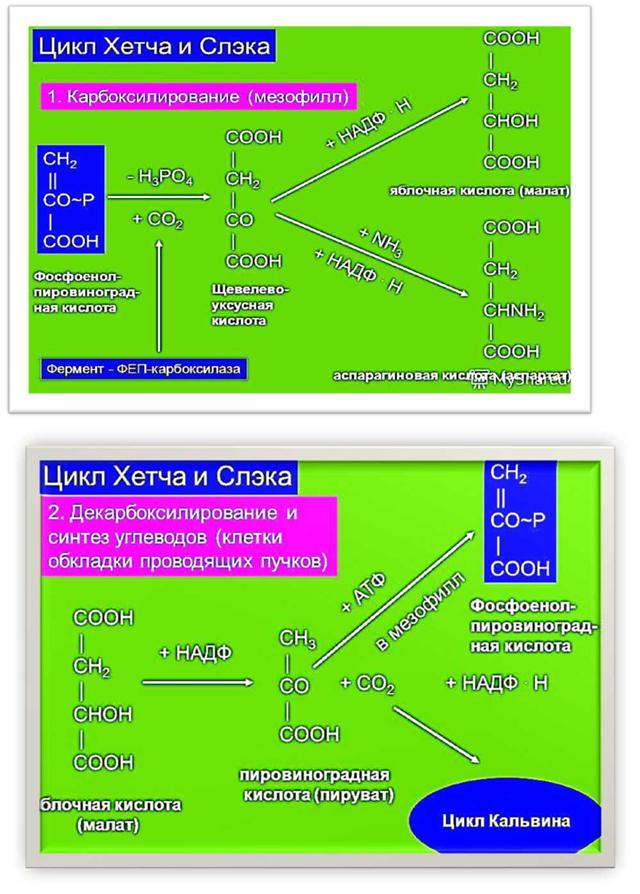 культурные сельскохозяйственные виды: просо, сорго, кукуруза, чумиза, сахарный тростник. В то же время среди С4- растений много злостных сорняков: пырей ползучий, марь белая, просо куриное, щетинник, щирица, речной гиацинт и другие виды. культурные сельскохозяйственные виды: просо, сорго, кукуруза, чумиза, сахарный тростник. В то же время среди С4- растений много злостных сорняков: пырей ползучий, марь белая, просо куриное, щетинник, щирица, речной гиацинт и другие виды.Исследования показали, что в растениях, в которых процесс фотосинтеза протекает по С4- пути, имеются два типа клеток и хлоропластов: мелкие гранальные пластиды в клетках мезофилла листа; крупные пластиды, часто лишенные гран, в клетках обкладки, окружающих сосудистые пучки. Клетки обкладки имеют утолщенные клеточные стенки, содержат большое количество хлоропластов и митохондрий, расположены вокруг сосудистых пучков в 1 или 2 слоя. Совокупность указанных особенностей анатомического строения получила название корончатой анатомии или корончатого синдрома (от слова kranz – корона). Хлоропласты разных типов клеток характеризуются не только особенностями строения, но и разным типам фосфорилирования. В клетках мезофилла по преимуществу происходит нециклическое фосфорилирование и образуется НАДФН, необходимый для цикла Кальвина, идущего в клетках обкладки. В хлоропластах клеток обкладки идет только циклическое фосфорилирование. Такое разделение типов фосфорилирования, возможно, связано с тем, что к хлоропластам клеток обкладки, расположенным в глубине листа, проникает по преимуществу более длинноволновый свет, который не поглощается фотосистемой, ответственной за разложение Н2О.Цикл Хэтча — Слэка также состоит из нескольких реакций, но он короче, чем С3-путь. Первая реакция этого цикла - фосфорилирование ПВК за счет АТФ и превращение ее в фосфоенолпируват (ФЕП). Далее происходит одновременно две реакции: карбоксилирование фосфоенолпировиноградной кислоты (присоединение двуокиси углерода), и ее дефосфорилирование - отщепление фосфорной кислоты. При этом образуется четырехчленное углеродное соединение – ЩУК: ФЕП + СО2 → ЩУК + Н3РО4. Это - первое четырехуглеродное соединение, продукт С4-пути фотосинтеза. Далее оно может реагировать различным образом: восстанавливаться с образованием яблочной оксикислоты (ЯК), или малата: ЩУК + 2Н — ЯК, либо аминироваться с образованием аспарагиновой аминокислоты (ААК): ЩУК + NH3 → ААК. При фотосинтезе по С4-пути непосредственно могут образоваться карбоновые кислоты и аминокислоты. Однако этот путь может быть связан с С3-путем. При этом яблочная кислота может вновь декарбоксилироваться, т. е. отщеплять углекислоту, а образующееся трехчленное углеродное соединение вступает в цикл Кальвина, который проходит уже не в столбчатой паренхиме, а в клетках обкладки проводящих пучков. В таком случае фотосинтез более активно идет при закрытых устьицах, так как углекислый газ запасается (образуется) в листе при более благоприятных условиях и пониженной транспирации. Доказательством более позднего появления этого пути считают наличие видов С 3- и С4- фотосинтеза в одних таксонах. С4-растения имеют физиологические преимущества, которые проявляются в условиях засухи, повышенной температуры, засоления почвы, сильного освещения, т. е. при условиях, характерных для более южных стран. У них меньше транспирационный коэффициент, следовательно, они тратят меньше воды на образование сухого вещества. Также меньше у них потребность в азоте. У этих растений отсутствует фотодыхание и наблюдается более высокая продуктивность фотосинтеза при повышенной температуре. 77. Характеристика фотосинтетического цикла Кальвина (С3-путь фотосинтеза). Первичный и конечный продукты фотосинтеза. Фотосинтез – синтез, совмещение, помещение вместе) – процесс образования органических веществ из углекислого газа и воды на свету при участии фотосинтетических пигментов (хлорофилл у растений, бактериохлорофилл и бактериородопсин у бактерий). В современной физиологии растений под фотосинтезом чаще понимается фотоавтотрофная функция – совокупность процессов поглощения, превращения и использования энергии квантов света в различных эндэргонических реакциях, в том числе превращения углекислого газа в органические вещества. В темновой стадии с участием АТФ и НАДФН происходит восстановление CO2 до глюкозы (С6Н12О6). Хотя свет не требуется для осуществления данного процесса, он участвует в его регуляции.  Единственным источником всех органических веществ, получающихся в процессе фотосинтеза, является углекислый газ СО2, который вступает в реакцию с водой. На самом деле должно пройти большое число реакций, прежде чем образуется, органическое вещество. Следовательно, фотосинтез представляет собой многоступенчатый процесс, который к тому же с трудом поддается исследованию. Сложность заключается в том, что первичные продукты фотосинтеза неустойчивы и подвержены быстрому превращению. Раскрытие химизма фотосинтеза стало возможным только тогда, когда были разработаны очень точные методы биохимического анализа, особенно хроматография и радиоизотопный метод с использованием изотопа углерода 14С, который в виде двуокиси углерода вводится в растение. Первым продуктом, который удалось зафиксировать при очень коротких световых экспозициях, было трехуглеродное соединение — фосфоглицериновая кислота (ФГК) СООН—СНОН— СН20(Р). Вследствие этого данный путь фотосинтеза, считающийся в настоящее время основным, носит название С3-пути. Единственным источником всех органических веществ, получающихся в процессе фотосинтеза, является углекислый газ СО2, который вступает в реакцию с водой. На самом деле должно пройти большое число реакций, прежде чем образуется, органическое вещество. Следовательно, фотосинтез представляет собой многоступенчатый процесс, который к тому же с трудом поддается исследованию. Сложность заключается в том, что первичные продукты фотосинтеза неустойчивы и подвержены быстрому превращению. Раскрытие химизма фотосинтеза стало возможным только тогда, когда были разработаны очень точные методы биохимического анализа, особенно хроматография и радиоизотопный метод с использованием изотопа углерода 14С, который в виде двуокиси углерода вводится в растение. Первым продуктом, который удалось зафиксировать при очень коротких световых экспозициях, было трехуглеродное соединение — фосфоглицериновая кислота (ФГК) СООН—СНОН— СН20(Р). Вследствие этого данный путь фотосинтеза, считающийся в настоящее время основным, носит название С3-пути.С3-фотосинтез, цикл Кальвина. Этот способ ассимиляции СО2, присущий всем растениям, был расшифрован американскими исследователями во главе с Мэльвином Кальвиным. Схема последовательности реакций по-лучила название цикла Кальвина. Характерной особенностью фотосинтетического восстановления СО2 являются цикличность и разветвленность этого процесса. Цикличность обеспечивает высокую производительность, саморегуляцию и непрерывность образования углеводов. Разветвленность - образование разнообразных продуктов, дублирование путей регенерации акцептора СО2. Цикл состоит из трех этапов: карбоксилирования, восста-новления и регенерации акцептора СО2Восстановительный пентозофосфатный цикл, цикл Кальвина – серия биохимических реакций, осуществляемая при фотосинтезе растениями (в строме хлоропластов), цианобактериями, прохлорофитами и пурпурными бактериями, а также многими бактериями- хемосинтетиками, является наиболее распространённым из механизмов автотрофной фиксации CO2. Цикл Кальвина или восстановительный пентозофосфатный цикл состоит из трёх стадий: карбоксилирования; восстановления; регенерация акцептора CO2. 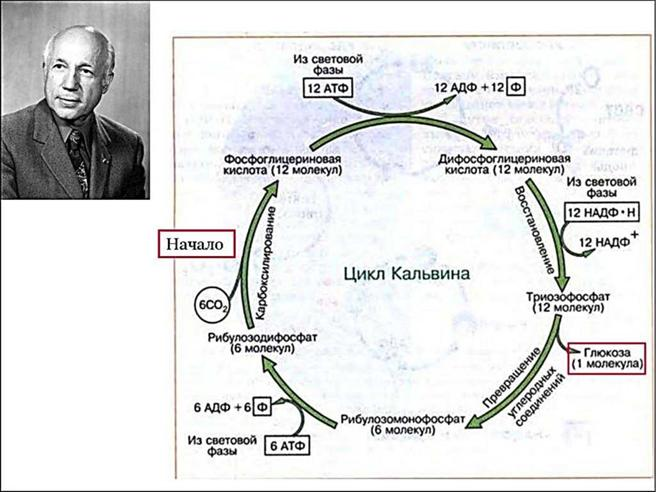 Карбоксилирование. Первой реакцией, вводящей СО2 в цикл Кальвина, является карбоксилирование рибулезо-1.5-дифосфата (1,5-РДФ) с участием фермента рибулезодифосфаткарбоксилазы (РДФ-карбоксилазы). Образующееся при этом нестойкое щести-утлеродное соединение быстро распадается на триозы - две молекулы 3- фосфоглицериновой кислоты (3-ФГК). Поэтому З-ФГК можно считать первичным продуктом фотосинтеза. На первой стадии к рибулозо-1,5-бифосфату присоединяется CO2 под действием фермента рибулозобисфосфат-карбоксилаза/оксигеназа (RuBisCO). Рибулозобисфосфат- карбоксилаза оксигеназа (Рубиско) - основной фермент цикла Кальвина, осуществляющий процесс фиксации углекислого газа в фотосинтезе. Рубиско состоит из восьми субъединиц: четырёх больших, осуществляющих каталитическую функцию и кодируемых в хлоропластном геноме, и четырёх малых, регулирующих работу фермента и кодируемых в ядре. Рубиско активируется на свету. В цикле Кальвина Рубиско присоединяет молекулу углекислого газа к пятиуглеродному сахару рибулозо-1,5-бисфосфату (РуБФ). Эта реакция называется карбоксилазной. Помимо карбоксилазной реакции, Рубиско способен катализировать оксигеназную реакцию-присоединять к РуБФ молекулу кислорода. Продукт этой реакции участвует в биохимическом цикле, называемом фотодыханием. Фотодыхание необходимо для синтеза некоторых аминокислот, а также снижения в клетке содержания кислорода, который в больших количествах может быть опасен. Рубиско составляет до 50% всего белка в листе и считается одним из самых распространённых ферментов биосферы. 1) Фаза восстановления. Восстановление 3-фосфоглицериновой кислоты (3-ФГК) до 3- фосфоглицеринового альдегида (3-ФГА) происходит в два этапа. Сначала при участии АТФ и фосфоглицераткиназы 3-ФГК присоединяет остаток фосфорной кислоты с образованием 1,3- дифосфоглицериновой кислоты (1,3-ДФГК.) Этим достигается повышение реакционной способности соединения и возможность его восстановления с помощью НАДФ- Н до З-ФГА. Это единственная восстановительная реакция цикла. Фаза восстановления является центральным звеном цикла. Именно здесь скрещиваются световая и темновая фазы фотосинтеза. Все остальные превращения идут на уровне сахаров, одинаковых по степени восстановленности. 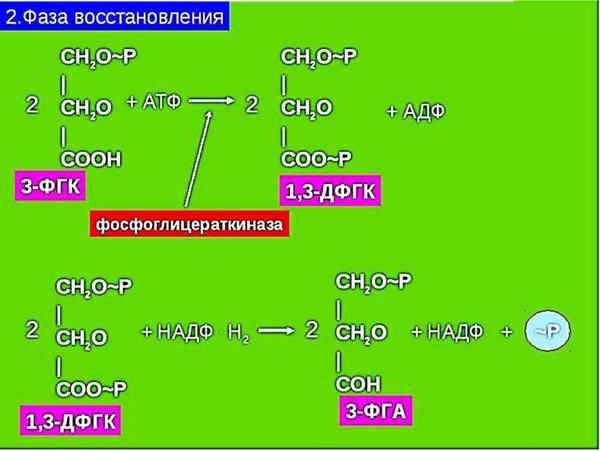 3) Фаза регенерации первичного акцептора СО2 и синтеза конечных продуктов фотосинтеза. В результате рассмотренных ранее реакций при фиксации трех молекул СО2 образуются 6 молекул восстановленных 3-фосфотриоз, пять из них используются затем для регенерации РДФ, а 1- для синтеза глю-козы. Это достигается следующей последовательностью реакций. Часть молекул З-ФГА под действиечтриозофосфатизомеразы превращается в фосфодиоксиацетон (ФДА). Затем два изомера (ФГА и ФДА) полвергаются конденсации при участии фермента альдолазы, образуя фруктоза-1,6-дифосфат, у которого затем от-щепляется один фосфат. 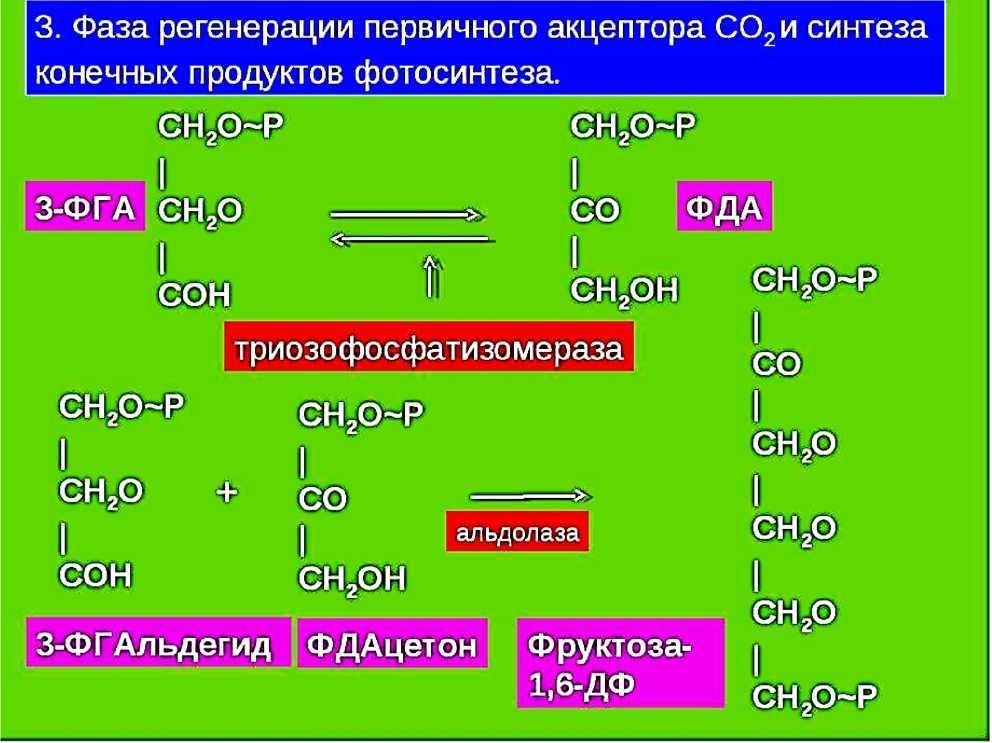 В дальнейших реакциях, связанных с регенерацией акцептора СО2, возникает цепь фосфорных эфиров сахаров, содержащих в своем составе 4, 5 или 7 атомов углерода. В этих реакциях последовательно принимают участие транскето-лазы и трансальдолазы. Транскетолаза катализирует перенос 2-угле-родной группировки от фруктоза-6-фосфата на З- ФГА, при этом образуются эритрозо-4-фосфат (С4) и ксилулозо-5-фосфат (С5). Затем альдолаза осуществляет перенос 3-углеродного остатка ФДА на эритрозо-4-фосфат, в результате чего синтезируется седо-гептулозо-1.7-дифосфат (С7). От последнего отщепляется один остаток фосфорной кислоты и под действием транскетолазы из него и З-ФГА образуются ксилулозо-5-фосфат (С5) и рибозо- 5--фосфат (С5). Две молекулы ксилулозо-5- фосфата путем эпимери-зации и одна молекула рибозо-5-фосфата за счет изомеризации превращаются в три молекулы рибулозо-5-фосфата (С5). Это соединение подвергается фосфорилированию за счет АТФ, образующейся в световую фазу. Из оставшейся неиспользованной молекулы ФГА при повторении цикла образуется фруктоза-1,6-дифосфат, из которой могут образовываться глюкоза, сахароза и крахмал. Таким образом, для синтеза одной молекулы ГЛЮКОЗЫ (С6) должно произойти шесть оборотов цикла. В каждом обороте используются 3 мол. АТФ (две для активирования двух молекул ФГК и одна при регенерации акцептора СО2 1,5--РДФ) и две молекулы НАДФН для восстановления ФГК до ФГА. Поэтому для синтеза одной молекулы глюкозы в цикле Кальвина необходимо 12 НАДФН и 18 АТФ. Таков энергетичес-кий вклад световой фазы в темновую фиксацию СО2. В цикле Кальвина первичными продуктами включення СО2 в органические вещества н восстановления являются трехуглерод-ные соединения (З-ФГК, З-ФГА, ФДА). Поэтому этот способ фиксации СО2 носит название С3-пути фотосинтеза. Большинст-во растений, особенно произрастающих в умеренной зоне, ис-пользуют именно этот путь восстановления СО2 и называются С3-растениями. 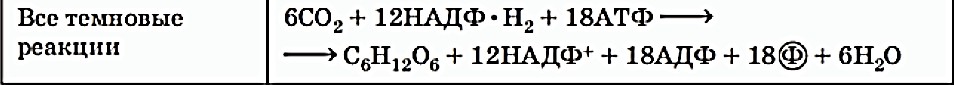 Суммарное уравнение всех темновых реакций Суммарное уравнение всех темновых реакций78. Характеристика активации кислорода и водорода и ее связь с процессом дыхания у растений. Теории А.Н.Баха и В.И.Палладина. Дыхание растительного организма (так же, как и животного) осуществляется в процессе биологического окисления. Но надо помнить, что окисления как такового в живой природе нет, оно непременно сопровождается восстановлением, поэтому правильнее говорить о едином о/в процессе. Биологическое окисление имеет несколько форм. Например, оно может происходить с потерей электрона. Возможна также потери водорода. Для понимания сущности о/в. процессов необходимо знание теорий биологического окисления. Первая до времени появления теория, разработанная русским биохимиком А.Н.Бахом (1897), носит название перекисной теории биологического окисления. Вторая теория была разработана В.И.Палладиным (1916). Теория активации кислорода (А. Н. Бах). Согласно этой теории непосредственным агентом, окисляющим субстрат является активированный кислород. Активация молекулярного кислорода осуществляется особыми ферментами оксигеназами и ведет к образованию реакционноспособных пероксидов, которые при участии ферментов пероксидаз и взаимодействуют с субстратом. 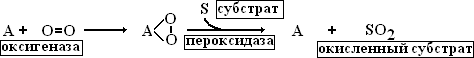 Основные положения: в окислении обязательно участвует кислород; при его активации разрывается только одна связь; О2 передается на субстрат через промежуточные вещества; участвуют ферменты – оксигеназы. Теория активации водорода (дегидрирование) (В. И. Палладин). По этой теории, биологическое окисление может происходить путем дегидрирования субстрата и без участия кислорода, но при помощи коферментов – акцепторов водорода, т.е. окисление на первых этапах может происходить и в анаэробных условиях, а затем может идти при участии кислорода в аэробных условиях, в результате чего образуются молекулы воды.  Предполагалось, что акцептором атомов водорода являются соединения хинонной при роды (подобные, убихинону). Сейчас установлено, что в процессе биологического окисления атомы водорода от субстрата отщепляются коферментами – НАД, НАДФ, ФАД и ФМН. В. И. Палладин впервые стал рассматривать дыхание как ряд ферментативных реакций Значение этой теории для последующего развития знаний по физиологии дыхания очень велико. В ней заложены основы современных представлений об этом процессе: дыхание как сложный о/в процесс; участие воды в дыхании как субстрата и продукта реакций; анаэробное окисление углевода и анаэробное происхождение углекислого газа; участие ферментов дегидрогеназ (прообразы их – гипотетические хромогены и пигменты) в процессе дыхания. Многие положения теории, впервые выдвинутые В. И. Палладиным, позднее были подтверждены другими исследователями. Символом R В. И. Палладии обозначал дыхательный пигмент, способный к обратимым окислительно-восстановительным превращениям. Непременным участником дыхания является вода. Вода наряду с окисляемым субстратом выполняет роль донора водорода. В процессе дыхания участвуют специфические активаторы водорода, отнимающие водород от субстрата. Первые этапы дыхания являются анаэробными и не требуют присутствия молекулярного кислорода. Молекулярный кислород используется на заключительном этапе дыхания для регенерации акцепторов водорода с образованием воды. 79. Теория генетическои связи процессов дыхания и брожения С.П.Костычева. Сергей Павлович Костычев доказал, что теория Пфеффера и Плюгера ( промежуточный продукт дыхания этанол) не действительна, так как этанол является ядом для растений и он плохо окисляется растительными тканями. Костычев установил настоящую генетическую связь между дыханием и брожением. В опытах Костычева и его сотрудников было показано, что если растительные ткани кратковременно выдержать в бескислородной среде, а затем дать кислород, то наблюдается резкое усиление дыхания, т.е. в ходе анаэробной фазы накапливаются промежуточные продукты, которые в присутствии кислорода быстро используются. Ингибиторы, блокирующие брожение, например фторид натрия, блокируют и аэробное дыхание. Ингибиторный анализ (применение ингибиторов специфического действия ), выделение и идентификация продуктов окислительного распада глюкозы привели Костычева к выводу о том, что промежуточным продуктом может быть уксусный альдегид. Благодаря этим работам стало очевидно, что дыхание и все виды брожения связаны между собой через пировиноградную кислоту (ПВК). Таким образом теория полностью подтвердилась. Гликолиз – процесс анаэробного распада глюкозы, идущий Гликолиз с освобождением энергии, конечным продуктом которого является пировиноградная кислота. Гликолиз осуществляется во всех живых клетках организма. В процессе гликолиза происходит преобразование молекулы гексозы до двух молекул пировиноградной кислоты. С.П. Костычев выдвинул положение о генетической связи процессов брожения и дыхания. Начальная фаза превращения сахара – анаэробный распад молекулы дыхательного субстрата – является общей и для дыхания, и для брожения. Через целую цепь последовательных превращений этот распад приводит к образованию промежуточных продуктов, опять-таки общих как для дыхания, так и для брожения. Пути этих двух про- цессов расходятся позднее, когда распад сахара достигает определенного этапа. Таким образом, устанавливается определенная связь между дыханием и брожением через пировиноградную кислоту (ПВК). |