ответы экзамен. Эволюция пищеварительной системы и особенности питания у представителей разных классов позвоночных животных
 Скачать 7.56 Mb. Скачать 7.56 Mb.
|

|
Содержание воды в разных органах растения:
В вакуолярном клеточном соке преобладает вода, удерживаемая сравнительно низкомолекулярными соединениями (осмотически-связанная) и свободная вода. В оболочке растительной клетки вода связана, главным образом, высокополимерными соединениями (целлюлозой, гемицеллюлозой, пектиновыми веществами), т. е. коллоидно-связанная вода. В самой цитоплазме имеется вода свободная, коллоидно- и осмотически-связанная. Вода, находящаяся на расстоянии до 1 нм от поверхности белковой молекулы, связана прочно и не имеет правильной гексагональной структуры (коллоидно-связанная вода). Кроме того, в цитоплазме имеется определенное количество ионов, а, следовательно, часть воды осмотически связана. Физиологическое значение свободной и связанной воды различно. Как считает большинство исследователей, интенсивность физиологических процессов, в том числе и темпов роста, зависит в первую очередь от содержания свободной воды. Имеется прямая корреляция между содержанием связанной воды и устойчивостью растений против неблагоприятных внешних условий. Указанные физиологические корреляции наблюдаются не всегда. Функции воды: Основные функции воды в растениях: Объединяет все части организма, образуя непрерывную водную фазу; Образует раствор и среду для реакций метаболизма; Принимает участие в различных процессах как вещество реакции 6СО2 + 6Н2О→С6Н12О6 + 6О2 Обеспечивает передвижение веществ по сосудам растения, по симпласту и апопласту; Защищает ткани растений от резких колебаний температуры (благодаря высокой теплоемкости и большой удельной теплоте парообразования); Обеспечивает упругость тканей и органов, выполняет роль амортизатора при механических воздействиях; Поддерживает структуру органических молекул, мембран, цитоплазмы, клеточной стенки и других компартментов клетки. Свойства воды, обеспечивающие ее функции в растительной клетке: молекула воды представляет собой диполь; благодаря этому молекулы воды могут ассоциировать друг с другом, ионами и белковыми молекулами; вода участвует в поглощении и транспорте веществ, так как является хорошим растворителем, гидратные оболочки, окружающие ионы, ограничивают их взаимодействие; вода обладает высокой теплоемкостью – 1кал/град, что позволяет растению воспринимать изменения температуры окружающей среды в смягченном виде, испарение воды растениями – транспирация служит основным средством терморегуляции у растений. Состояние и фракционный состав внутреклеточной воды В клетках и тканях различают две формы воды – прочно связанную (связанную) и рыхлосвязанную (свободную). Связанная вода. Связанная вода – имеет измененные физические свойства главным образом в результате взаимодействия с неводными компонентами. Условно принимают под связанной водой ту, которая не замерзает при понижении температуры до – 10 °С. Связанная вода в растениях бывает: Осмотически- связанная Коллоидно-связанная Капиллярно-связанная Осмотически-связанная вода – связана с ионами или низкомолекулярными веществами. Вода гидратирует растворенные вещества – ионы, молекулы. Вода электростатически связывается и образует мономолекулярный слой первичной гидратации. Вакуолярный сок содержит сахара, органические кислоты и их соли, неорганические катионы и анионы. Эти вещества удерживают воду осмотически. Коллоидно-связанная вода – включает воду, которая находится внутри коллоидной системы и воду, которая находится на поверхности коллоидов и между ними, а также иммобилизованную воду. Иммобилизация представляет собой механический захват воды при конформационных изменениях макромолекул или их комплексов, при этом вода оказывается заключенной в замкнутом пространстве макромолекулы. Значительное количество коллоидно-связанной воды находится на поверхности фибрилл клеточной стенки, а также в биоколлоидах цитоплазмы и матриксе мембранных структур клетки. Осмотически-связанная вода – гидратирует растворенные вещества – ионы и молекулы; коллоидно-связанная вода ‒ гидратирует коллоиды (макромолекулы); капиллярно-связанная вода – связанная со структурами клеточных стенок и сосудов за счет сил адгезии. Связанная вода выполняет структурную функцию – поддерживая структуру коллоидов и обеспечивая функционирование ферментов, органоидов и клетки в целом. Она мало подвижна, не участвует в растворении и транспорте веществ, отличается более низкой температурой замерзания и более высокой температурой кипения, чем свободная вода. Свободная вода обладает высокой подвижностью, является растворителем и основным транспортером веществ по растению. Доля связанной воды в клетке составляет около 40 %, доля свободной – около 60 %. При недостатке влаги в первую очередь снижается доля свободной воды. Структура воды (рис. 2.1): атом О обладает сильной электроотрицательностью и стремиться оттянуть электроны от атомов Н: на атоме О – частично отрицательный заряд (–), 2 атома Н приобретают частично положительный заряд (+); несущие положительный заряд атомы Н испытывают электростатическое притяжение со стороны отрицательно заряженных атомов О соседних молекул Н2О; малые размеры молекул; полярность молекул – неравномерное распределение зарядов в молекуле: один конец молекулы несет небольшой положительный заряд, а другой – отрицательный: молекула Н2О – диполь (полярная молекула) (рис.2.1); способность молекул соединяться друг с другом водородными связями (энергия около 4,8 ккал/моль); 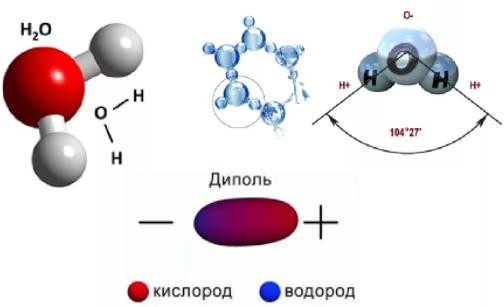 большая упорядоченность молекул в водных растворах. большая упорядоченность молекул в водных растворах.Рисунок 2.1‒ Структура воды 72. Общая характеристика пигментов, участвующих в процессе фотосинтеза. ПИГМЕНТЫ – природные вещества, избирательно поглощающие свет в видимом участке спектра. Основное назначение пигментов – поглощать световую энергию, превращая ее затем в химическую энергию. Пигменты располагаются на мембранах хлоропластов (тилакоидах), а хлоропласты в клетке обычно ориентируются таким образом, чтобы мембраны находились под прямым углом к источнику света (для максимального поглощения света). Хлорофиллы: хлорофилл а – у большинства фотосинтезирующих организмов, хлорофилл b – у высших растений и зелёных водорослей, хлорофилл с – у бурых водорослей, хлорофилл d – у некоторых красных водорослей. Каротиноиды: каротины –у всех фотосинтезирующих организмов, кроме прокариот; ксантофиллы –у всех фотосинтезирующих организмов, кроме прокариот Фикобилины – красные и синие пигменты красных водорослей. В хлоропластах пигменты ассоциированы с белками с помощью ионных, водородных и других типов связей. Хлорофиллы. В настоящее время известно около десяти хлорофиллов. Они отличаются по химическому строению, окраске, распространению среди живых организмов. У всех высших растений содержатся хлорофиллы а и b. Хлорофилл с обнаружен в диатомовых водорослях, хлорофилл d – в красных водорослях. Кроме того, известны четыре бактериохлорофилла (a, b, с и d), содержащиеся в клетках фотосинтезирующих бактерий. В клетках зеленых бактерий имеются бактериохлорофиллы с и d, в клетках пурпурных бактерий – бактериохлорофиллы а и b. Основными пигментами, без которых фотосинтез не идет, являются хлорофилл а для зеленых растений и бактериохлорофиллы для бактерий. Впервые точное представление о пигментах зеленого листа высших растений было получено благодаря работам крупнейшего русского ботаника М. С. Цвета (1872—1919). Хлорофилл b отличается от хлорофилла а тем, что у 3-го углерода вместо метильной находится формильная (—СНО) группа. Структура хлорофилла, лишенная фитола, называется хлорофиллидом. При замещении атома магния протонами в молекуле хлорофиллов образуются соответствующие феофетины. Хлорофилл – это зелёный пигмент, обуславливающий окраску зелёного цвета растению, при его участии обусловлен процесс фотосинтеза. По химическому строению это Mg-комплекс различных тетрапирролов. Хлорофиллы имеют порфириновое строение, структурно близки к гему. В пиррольных группировках хлорофилла имеются системы, чередующихся двойных и простых связей. Это и есть хромофорная группа хлорофилла, обуславливающиеся поглощение определённых лучей солнечного спектра и его окраску. D порфировые ядра составляют 10 нм, а длина фитольного остатка 2 нм. Молекулы хлорофилла полярно, её порфириновое ядро обладает гидрофильными свойствами, а фитольный конец гидрофобными. Это свойство молекулы хлорофилла обуславливают определённое расположение её в мембранах хлоропласта. Порфириновая часть молекулы связана с белком, а фитольная часть погружена в липидный слой. Хлорофилл живой интактной клетки обладает способностью к обратимому фотоокислению и фотовосстановлению. Способность к окислительно-восстановительным реакциям связано с наличием в молекуле хлорофилла сопряжённых двойных связей с подвижными п-элктронами и атомами N с неопределёнными электронами. ФИЗИОЛОГИЧЕСКАЯ РОЛЬ: избирательно поглощать энергию света, запасать ее в виде энергии электронного возбуждения, фотохимически преобразовывать энергию возбужденного состояния в химическую энергию первичных фотовосстановленных и фотоокисленных соединений. Каротиноиды– это жирорастворимые пигменты желтого, оранжевого, красного цвета присутствуют в хлоропластах всех растений. Каротиноиды содержатся во всех высших растениях и у многих микроорганизмов. Это самые распространенные пигменты с разнообразными функциями. Каротиноиды имеют максимальное поглощение в фиолетово- синей и синей частях спектра света. Они не способны к флуоресценции в отличие от хлорофилла. К каротиноидам относятся 3 группы соединения: оранжевые, или красные каротины; жёлтые ксантофиллы; каротиноидные кислоты. ФИЗИОЛОГИЧЕСКАЯ РОЛЬ: Поглощение света в качестве дополнительных пигментов; Защита молекул хлорофилла от необратимого фотоокисления; Тушение активных радикалов; Участвуют в фототропизме, т.к. способствуют направлению роста побега. Фикобилины – это красные и синие пигменты, содержащиеся у цианобактерий и некоторых водорослей. Фикобилины состоят из 4-х последовательных пиррольных колец. Фикобилины являются хромофорными группами глобулиновых белков, который называется фикобилинпротеинами. Он делятся на: -фикоэритрины – белки красного цвета; -фикоцианин – синеголубые белки; алофикоцианин – синие белки. Все они обладают флуоресценирущей способностью. Фикобилины имею максимальное поглощение в оранжевых, жёлтых и зелёных частях спектра света и позволяют водорослям полнее использовать свет, проникающий в воду. На глубине 30 м полностью исчезают красные лучи На глубине 180 м – жёлтые На глубине 320 м – зелёные На глубине более 500 м не проникают синие и фиолетовые лучи. Фикобилины - это дополнительные пигменты примерно 90% энергии света, поглощающего фикобилинами передаётся на хлорофилл. ФИЗИОЛОГИЧЕСКАЯ РОЛЬ: Максимумы поглощения света у фикобилинов находятся между двумя максимумами поглощения у хлорофилла: в оранжевой, желтой и зеленой частях спектр. Фикобилины выполняют у водорослей функции светособирающего комплекса. У растений имеется фикобилин-фитохрм, он не участвет в фотосинтезе, но является фоторецептором красного света и выполняет регуляторную функцию в клетках растений. 73. Этапы гликолиза в растительной клетке и его функции. Гликолиз – последовательность реакций, в ходе которых глюкоза последовательно окисляется до пирувата. Самый древний энергетический путь. Выход: 2 молекулы АТФ, 2 молекулы НАДН. У растений гликолиз идет в двух компартментах – в цитозоле и в пластидах. В пластидах – возможно, с другими задачами, в хлоропластах – как правило в темноте. Исходное соединение для гликолиза у растений - сахароза, а не глюкоза, как у животных Субстрат для гликолиза – глюкоза. Глюкоза образуется в клетке при гидролизе 1)крахмала или 2) сахарозы. При гликолизе одна молекула глюкозы расщепляется до двух молекул пировиноградной кислоты (ПВК). Из АДФ синтезируется АТФ. Однако процесс идет с небольшим выделением энергии (1 М глюкозы – 200 кДж) Существует три этапа гликолиза: 1 – подготовительный (фосфорилирование гексозы и образование двух фосфотриоз); 2 – первое окислительное субстратное фосфорилирование;  3 – второе внутримолекулярное окислительное субстратное фосфорилирование. 1. Образование фруктозо-1,6-бисфосфата Субстратом для начальных стадий гликолиза могут являться либо глюкоза, либо фруктоза, главное на выходе получить фруктозо-1,6-бисфосфат Путь в 3 реакции, из глюкозы. 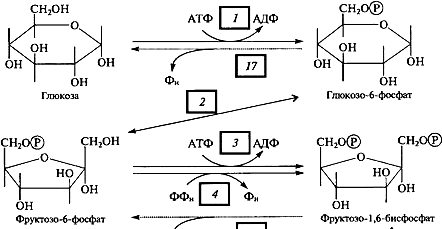 1 фермент – гексокиназа, 2 фермент – фосфоглюкоизомераза, 3 фермент – АТФзависимая фосфофруктокиназа , 4 – пирофосфатзависимая фосфофруктокиназа (катализирует реакцию в обоих направлениях). Обратите внимание, что последняя стадия может идти 2 способами, с участием 2 разных ферментов. Пирофосфат как субстрат для фосфорилирования больше никто, кроме растений не использует. У животных он быстро расщепляется в цитозоле, а у растений нет такого фермента. Путь, из фруктозы. По сути сокращенный вариант первого пути. Просто получаем фруктозо-6-фосфат непосредственно из фруктозы. Фермент, фосфорилирирующий фруктозу до фруктозо-6-фосфата – тоже гексокиназа. 3 Путь, нестандартный. Из дисахарида сахарозы. → Сахароза UDP-глюкоза, с участием фермента сазарозсинтазы. UDP – уридиндифосфат. → UDP-глюкоза глюкозо-1-фосфат, фермент – UDP-глюкозопирофосфорилаза, использует в качетстве субстрата пирофосфат. → глюкозо-1-фосфат глюкозо-6-фосфат, фермент - фосфоглюкомутаза. далее аналогично первому пути. 2. Распад фруктозо-1,6-бисфосфата С участием фермента альдолазы до глицеральдегид-3-фосфата(3ФГА) и фосфодиоксиацентона(ФДА), который легко изомеризуется до 3ФГА.  Окислительно-восстановительные реакции, тут образуется АТФ и NADH. II.Субстратное фосфорилирование (реакции 1-2) Окисление 3ФГА. Фермент глицеральдегид-3-фосфат-дегидрогеназа. Образуется НАДН. Продукт – дифосфоглицерат – макроэргическое соединение. 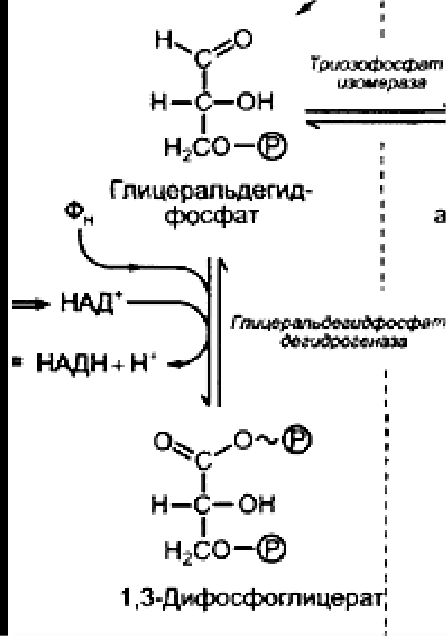 Перенос макроэргического Ф на АДФ. Фермент – фосфоглицераткиназа. Результат – образование первой АТФ. 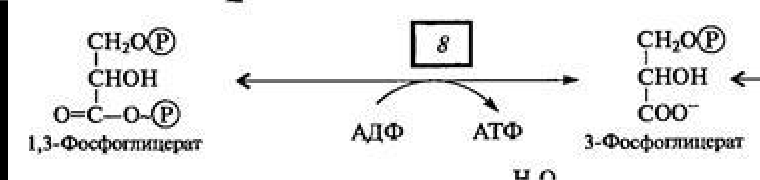 *** у растений есть НАДФ-зависимая-глицеральдегид-3-фосфат дегидрогеназа, которая катализирует реакцию (при фосфорном голодании) 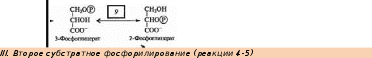 |