ответы экзамен. Эволюция пищеварительной системы и особенности питания у представителей разных классов позвоночных животных
 Скачать 7.56 Mb. Скачать 7.56 Mb.
|

|
Отщепление воды, фермент – енолаза. Образование высокоэнергетического соединения – фосфоенолпирувата (ФЕП) 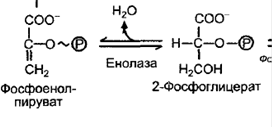 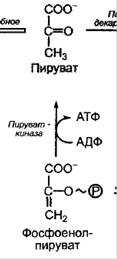 1. Перенос Ф на АДФ, фермент – пируваткиназа. Образованный пируват – конечный продукт гликолиза. Конечный продукт гликолиза у растений - не только пируват, (пировиноградная кислота) но и малат (яблочная кислота) *** у некоторых растений фермент ФЕП-фосфатаза, катализирует реакцию (при фосфорном голодании)  ФЕП-карбоксилаза катализирует синтез ООА из ФЕП Малик-энзим катализирует окислительное декарбоксилирование малата Смысл: эта реакция позволяет обходить последнюю стадию гликолиза (образование пирувата из ФЕП) и экономить ФЕП для цикла Кребса. ФЕП – ООА – малат, а малат может встраиваться в ЦТК (если есть такая необходимость) или окисляться до пирувата. 6. Шунтирование – пентозофосфатный окислительный цикл.  Регуляция гликолиза Нереализованный ФЕП и АТФ тормозит гликолиз, а АДФ, ионы фосфата, фруктозо- 2,6-бисфосфат стимулируют процесс. Пируваткиназа аллостерически ингибируется метаболитами ЦТК, активируется АДФ и фосфатом. Особенности гликолиза в растительной клетке: Проходит не только в цитозоле, но возможен и в хлоропластах в темноте (т.е. есть разные изоформы ферментов, которые катализируют реакции гликолиза; пластидные и цитозольные изоформы кодируются разными ядерными генами; пирофосфатзависимая фосфофруктокиназа не встречается в хлоропластах). Растения используют ферменты, использующие макроэргические связи пирофосфата. Продукты реакций гликолиза могут пополнять цикл Кребса (реакции, которые позволяют пополнять пул другого процесса называются анаплеротическими): из ФЕП образуется оксалоацетат (ООА), фермент ФЕП-карбоксикиназа; из пирувата образуется ООА, фермент пируваткарбоксилаза; из ООА образуется малат (обратимая реакция), фермент НАД-зависимая малатдегидрогеназа. Энергетический выход гликолиза  Функции гликолиза в клетке. В аэробных условиях гликолиз выполняет ряд функций: осуществляет связь между дыхательными субстратами и циклом Кребса; поставляет на нужды клетки две молекулы АТФ и две молекулы НАДН при окислении каждой молекулы глюкозы (в условиях аноксии гликолиз, по-видимому, служит основным источником АТФ в клетке); производит интермедиа, необходимые для синтетических процессов в клетке (например, фосфоенолпируват, необходимый для образования фенольных соединений и лигнина); в хлоропластах гликолитические реакции обеспечивают прямой путь для синтеза АТФ, независимый от поставок НАДФН 74. Механизмы регуляции устьичной транспирации. Устьица – это отверстия в эпидермисе, через которые происходит газообмен. Они находятся в основном на листьях, но имеются также и на стеблях. Каждое устьице окружено двумя замыкающими клетками, которые в отличие от обычных эпидермальных клеток содержат хлоропласты. Замыкающие клетки контролируют величину отверстия устьица за счет изменения своей тургесцентности. Стенки замыкающих клеток неравномерно утолщены: стенка, которая ближе к отверстию устьица, называемая вентральной, толще, чем противоположная, называемая дорсальной. Кроме того, целлюлозные микрофибриллы в стенке ориентированы таким образом, что вентральная стенка менее эластична, чем дорсальная. 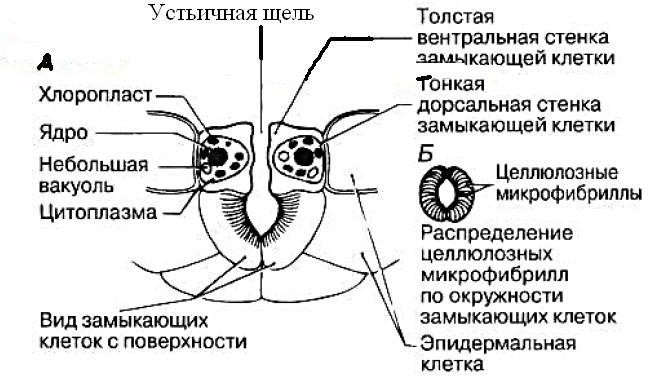 Скорость транспирации зависит, прежде всего, от степени открытости устьиц. Устьице открывается и закрывается при изменении тургорного давления в замыкающих клетках, что зависит от количества воды в замыкающих клетках. Устьице открывается, если в замыкающие клетки поступает вода, что приводит к увеличению тургорного давления в этих клетках и изменению их формы. Если из замыкающих клеток вода уходит, тургорное давление в них уменьшается, форма клеток изменяется, и устьице закрывается Существует несколько механизмов, влияющих на изменение тургорного давления в замыкающих клетках. Выделяют 2 механизма регулировки устьичных движений: устьичная регулировка; внеустьичная регулировка. Устьичная регулировка фотактивная реакция – под действием света устьица открываются; При хорошем освещении, чем выше уровень освещенности, тем шире открываются устьица. Установлено, что днем на свету в замыкающих клетках увеличивается содержание сахаров и, соответственно, повышается осмотическое давление. В замыкающих клетках увеличивается сосущая сила и в них поступает вода. Это приводит к открыванию устьиц. Кроме того, днем на свету взамыкающих клетках накапливаются и ионы калия, что также приводит к повышению осмотического давления и открыванию устьиц. (В ответ на освещение начинается фотосинтез в замыкающих клетках, и концентрация СО2 в межклетниках падает, поэтому устьице открывается. Увеличение устьичной щели уменьшает сопротивление диффузии СО2, его концентрация в межклетниках увеличивается, и устьице закрывается). гидроактивная (гидродинамичная)реакция закрывания устьиц – при сильной потере воды устьицами, реакция закрывания превышает фотоактивную реакцию закрывания: Гидродинамический механизм – это гидропассивная реакция, т.е. не связанная с изменением величины осмотического потенциала. Гидроактивное закрывание устьиц происходит в результате того, что транспирация превышает поглощение воды корнем. При этом тургорное давление замыкающих клетках достигает критического уровня. По мере развития водного дефицита в клетках листа увеличивается содержание фитогормона абсцизовой кислоты (АБК), которое приводит к снижению тургора клеток, и устьица закрываются. Вероятно, гидродинамический механизм работает, когда нарушается поступление воды в листовую пластинку. Например, если корни переносят в питательный раствор с пониженной температурой или перерезают черешок листа, то уже через несколько секунд увеличивается ширина устьичной щели. У срезанных веток кутикулярная транспирация идет быстрее, чем устьичная; примыкающие клетки теряют воду, и устьица открываются. У гидропассивная реакция закрывания устьиц – это случай, когда лист пренасыщен водой, его клетки приобретают сильное тургорное давление, давят на устьица и замыкают их. Гидропассивная устьичная реакция происходит тогда, когда клетки эпидермиса и хлоренхимы, окружающие устьица, после обильного полива переполняются водой и механически сжимают замыкающие клетки. Если окружающие клетки теряют воду, то устьица открываются. Внеустьичная регулировка Лист теряет много воды и при широко окрытых устьицах может прекратить транспирацию, так как коллоиды протоплазмы удерживают воду. Примеры. У хлебных злаков устьица всегда закрыты ночью, днем, при достаточном увлажнении, устьица открыты. Обычно с 5 часов утра до 11 часов дня. У тонколистных мезофитов (горох, донник, клевер, фасоль, репа, люцерна) при благоприятных условиях устьица открыты днем и закрыты ночью. Открываются через 2-3 часа после восхода солнца и закрываются постепенно после обеда, примерно в 2 часа дня. У растений с толстыми листьями (капуста, картофель) при высокой влажности устьица открыты и днем и ночью. В XIX в. движения устьиц связывали с хорошо известной уже тогда реакцией превращения крахмала в глюкозо-1-фосфат. Реакция обратима и катализируется крахмал-фосфорилазой, широко распространенной в листьях. Крахмал, в отличие от глюкозо-1-фосфата, не растворяется в воде, поэтому его концентрация не влияет на величину водного потенциала замыкающих клеток. Если крахмал превращается в глюкозо-фосфат, то уменьшается водный потенциал, вода поступает в замыкающие клетки, и устьице открывается. Если, наоборот, глюкозо- фосфат превращается в крахмал, то водный потенциал увеличивается, и вода уходит из замыкающих клеток, устьице закрывается. В результате увеличения концентрации глюкозо-фосфата осмотический потенциал может уменьшиться с -1 ... -2 до -10 МПа. Связь этой реакции с движениями устьиц получила название осмотического механизма. Превращение крахмала в сахар и обратно связано с ферментами, активность которых зависит от величины pH. Выкачивание Н+ из цитоплазмы в анонласт приводит к увеличению pH в замыкающей клетке и превращению крахмала в сахар, уменьшение pH вызывает противоположную реакцию. Другой механизм, изученный в XX в., был назван калиевым насосом. Тургорное давление зависит от перераспределения ионов калия между замыкающими и примыкающими клетками эпидермы. Ионы калия являются главным осмотиком растительных, в том числе и замыкающих, клеток. Поскольку замыкающие клетки не имеют плазмодесм, все ионы должны идти в цитозоль и из него - только через плазмалемму. В ответ на изменение внешних или внутренних условий на плазмалемме замыкающих клеток начинает работать Н+-помпа, откачивающая протоны из этих клеток в апопласт (1 Н+ на одну гидролизованную молекулу АТФ). Поставщиком АТФ является дыхание или фотосинтез в замыкающих клетках. В результате происходит гиперноляризация нлазмалеммы замыкающих клеток (более 100 мВ), и на ней открываются входные потенциалзависимые К+- каналы, через которые в вакуоль поступают ионы калия. Степень открытости этих каналов значительно увеличивается при большой разности электрического мембранного потенциала. В одном и том же листе степень открытости устьиц может быть разной. Доказательством выхода протонов из замыкающих клеток служит повышение величины pH при поступлении в эти клетки ионов калия. Источником протонов могут быть органические кислоты клеточного сока (малат); при открывании устьиц их содержание увеличивается. Чтобы при поступлении калия в клетки в них не изменился электрический заряд, одновременно в замыкающие клетки входят анионы. У двудольных растений противоионом служит в основном анион малата, у однодольных – С1 -. Анион малата накапливается в вакуоли вместе с калием. Малат в замыкающих клетках образуется из накопленного в хлоропластах крахмала, который в результате гидролиза и гликолиза превращается в фосфоенолпируват (ФЕП). ФЕП реагирует с НСО3 с образованием оксалоацетата. Реакцию катализирует ФЕП-карбоксилаза. ФЕП- карбоксилазу активирует увеличение концентрации ионов калия и подщелачивание цитозоля из-за выхода Н+. Оксалоацетат восстанавливается до малата с помощью НАДФ- малатдегидрогеназы. Закрывание устьиц происходит в результате обратного хода этих процессов. При закрывании устьиц основная роль в передаче сигнала и регуляции работы каналов принадлежит ионам Са2+, которые деполяризуют мембрану. Деполяризация мембраны (потенциал становится более положительным) вызывает открывание выходных К+-каналов. Ионы К+ выходят из вакуоли и идут в апопласт. Водный потенциал замыкающих клеток увеличивается, вода из этих клеток уходит, и устьице закрывается. Открытие ионных каналов может регулировать также фосфорилирование пептидов, образующих канал. При закрывании устьиц большая часть малата выходит из замыкающих клеток. Таким образом, движения устьиц связаны с процессами, происходящими на плазмалемме, в цитозоле, гонопласте и вакуоли замыкающих клеток, затрагивают апопласт и соседние клетки мезофилла. Ширина устьичной щели зависит от тургора не только в замыкающих, но и в примыкающих клетках эпидермы. Если в примыкающих клетках много воды, например сразу после дождя, то они давят на замыкающие клетки, и устьице не может открыться. Поэтому в дождливую влажную погоду, несмотря на высокое тургорное давление в замыкающих клетках, устьице остается закрытым. В условиях засухи первыми теряют воду примыкающие клетки, имеющие более тонкие стенки. Их объем уменьшается. В результате давление на замыкающие клетки исчезает, и устьице открывается. Таким образом, первой реакцией на засуху является усиление транспирации, что способствует увеличению поступления воды в корень. Затем устьице закрывается, так как за примыкающими теряют тургор и сами замыкающие клетки. Этот механизм открывания устьиц получил название гидродинамического. Использование разных механизмов для выполнения одной функции – один из главных принципов организации и функционирования клетки, который, используя кибернетические термины, можно назвать принципом надежности', если один механизм не может работать, то работает другой, функция продолжает осуществляться. Кроме того, из сочетания различных механизмов можно построить много разных систем, а многообразие всегда ведет к лучшему приспособлению. Разные механизмы взаимодействуют друг с другом. Взаимодействие фотосинтетического механизма и калиевых насосов видно из приведенного выше объяснения. Известно, что активность крахмал-фосфорилазы зависит от величины pH. Во время работы калиевых насосов из замыкающих клеток уходят протоны, что вызывает увеличение pH и, следовательно, приводит к активации этого фермента. В результате крахмал превращается в глюкозо-фосфат, понижается водный потенциал, вода поступает в замыкающие клетки, и устьице открывается. Следовательно, в данном случае взаимодействуют калиевые насосы и осмотический механизм устьичных движений. Движения устьиц зависят от изменения внешних условий и состояния клеток, органов, организма. 75. Этапы онтогенеза высших растений. Онтогенезом называют индивидуальное развитие организма от зиготы или вегетативного зачатка до естественной смерти. В ходе онтогенеза реализуется наследственная информация организма - его генотип - в конкретных условиях окружающей среды, в результате чего формируется фенотип, то есть совокупность всех признаков и свойств данного индивидуального организма. Онтогенез включает в себя все жизненные процессы и продолжается у разных растений от 10-14 дней до 3-5 тыс. лет. По продолжительности жизни растения делят на эфемеры, однолетние, двулетние и многолетние (секвойи — до 5 тыс. лет). Независимо от продолжительности жизни все растения можно разделить на две группы: моно- и поликарпические. Монокарпическими (греч. mono - один, karpos - плод) называются растения, цветущие и плодоносящие один раз в своей жизни (эфемеры, однолетние растения, двулетние растения (мокровь, свекла) и некоторые многолетние, например мексиканские агавы, бамбук) Наступление плодоношения у таких организмов приводит к их быстрому старению и отмиранию. Поликарпические растения - это растения, плодоносящие много раз в жизни. Онтогенез дискретен, т. е. его можно разделить на отдельные этапы, проходящие последовательно один за другим. В онтогенезе цветковых растений выделяют 5 этапов: эмбриональный, ювенильный (молодости), зрелости, размножения (генеративный) и старости (сенильный). Эмбриональный этап онтогенеза семенных растений охватывает развитие зародыша от зиготы до созревания семени включительно. В начале своего развития зародыши зависят от фитогормонов, поступающих из других тканей, прежде всего из эндосперма. Затем они сами начинают синтезировать гормоны, а при формировании семян ауксин выделяется из них в окружающие ткани, активируя рост околоплодника. На последнем этапе созревания семена теряют значительное количество воды и переходят в состояние покоя (у большинства видов средней полосы). Этот переход связан с уменьшением в тканях свободных ауксинов, цитокининов и гиббереллинов и с увеличением АБК. Ювенильный этап (молодость) у семенных растений начинается с прорастания семян или органов вегетативного размножения (например, клубней) и характеризуется быстрым накоплением вегетативной массы. Растения в этот период не способны к вегетативному размножению. В ювенильный период осуществляется прорастание семян (или вегетативных зачатков) и формирование вегетативных органов. Прорастание, в свою очередь, делится на фазы набухания семян, проклевывания, гетеротрофного роста проростка, перехода к автотрофному способу питания. Продолжительность ювенильного периода у различных растений неодинакова: от нескольких недель (однолетние травы) до десятков лет (у древесных). Проростки по многим параметрам не похожи на взрослые растения. Различия чаще всего заметны в форме листьев. У некоторых растений наблюдаются различия во внутреннем строении. Этап характеризуется полным отсутствием цветения или цветение слабо выражено даже при самых благоприятных для этого условиях. У растений проявляется большая способность к корнеобразованию. Ювенильное состояние поддерживается определенным соотношением гормонов. Этап зрелости и размножения т.е. период готовности к зацветанию и образованию органов вегетативного размножения, период закладки и роста органов размножения, формирования семян и плодов. В этот период растение наиболее жизнеспособно, оно сформировало вегетативную массу, достаточную для обеспечения роста и развития цветков, семян и плодов, которые являются гетеротрофными органами. Этап старости и отмирания. Этап старости и отмирания включает в себя период от полного прекращения плодоношения до естественной смерти организма. Это период прогрессирующего ослабления жизнедеятельности. Продолжительность жизни растений, принадлежащих к различным таксономическим группам и различным жизненным формам, варьирует в очень широких пределах: Старение и смерть - завершающие фазы онтогенеза любого многоклеточного растения, но термин «старение» может быть применен не только к целому растению, но и к отдельным органам - листьям, плодам, частям цветков. Старение имеет большое биологическое значение. Оно является одним из способов адаптации растений к неблагоприятным условиям среды. Кроме того, старение, по-видимому, способствует более быстрой эволюции, т.к. ускоряет смену поколений, т.е. «оборачиваемость» генетического материала. |