ответы экзамен. Эволюция пищеварительной системы и особенности питания у представителей разных классов позвоночных животных
 Скачать 7.56 Mb. Скачать 7.56 Mb.
|

|
Химическая структура липидов на примере фосфолипидов Молекулы фосфолипидов – сложные эфиры трехатомного спирта глицерина и остатка жирных кислот с длинными углеводородными цепями. Фосфолипиды имеют полярную гидрофильную голову – производные фосфатидной кислоты. Голова хорошо взаимодействует с полярными растворителями (водой), заряжена отрицательно. Выделяют два гидрофобных углеводородных хвоста, которые не взаимодействуют с водой. Один из хвостов, как правило, имеет 1 или больше двойных связей (ненасыщенная жирная кислота), у другого хвоста двойных связей нет (насыщенная жирная кислота). Второй основной функциональный компонент биологической мембраны – белки. Химические свойства мембранных белков, их структура и функции недостаточно изучены, т.к. они в биологической мембране находятся в контакте с липидами и их трудно выделить в чистом виде, не нарушая их структуру. Как правило, нерастворимы в воде и плохо растворимы в органическом растворителе. Одни из белков прочно связаны с биологической мембраной, другие легко отделяются при обработке водно-солевыми растворами: внутренние (структурные) и наружные (периферические) белки. 1902 г.: мембраны состоят из тонкого слоя фосфолипидов. На разделе воздух-вода фосфолипиды образуют молекулярный слой, при этом их головы находятся в полярной среде (вода), а неполярный хвост ориентирован в неполярную среду (воздух). Поэтому предположили, что биологические мембраны построены из монослоя липидов. 1925 г.: Гортер и Грендель показал, что площадь монослоя липидов, выделенных из мембран эритроцитов в 2 раза больше суммарной площади эритроцита. На основании этих результатов высказали идею, что липиды в мембране расположены в виде бимолекулярного слоя. Эту гипотезу подтвердили исследования электрических параметров биологической мембраны, а именно это высокие электрические сопротивления и большая ее электрическая емкость, т.е. биологические мембраны можно рассмотреть как электрический конденсатор. 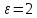  В этом конденсаторе пластинами являются электролиты, как с наружными, так и со внутренними растворами с погруженными в них головами фосфолипидов. Эти проводники (электролит) разделен диэлектрическим слоем, образованным неполярной частью липидных молекул, т.е. двойным слоем их хвостов. Липиды – диэлектрики с диэлектрической проницаемостью .Имелись данные, которые свидетельствуют, что биологические мембраны состоят не только из липидных молекул, но и из белковых молекул. Обнаружено, что измерение значения коэффициента поверхностного натяжения на границе раздела белок-вода ближе к такому коэффициенту, чем на границе раздела липиды-вода. Такие противоречивые результаты были установлены Даниэли и Даусоном в 1935 году, которые предположили т.н. «бутербродную модель» или «сендвича», согласно которой мембрана трехслойная т.е. она образована двумя слоями расположенных по краю молекул белка и липидным слоем по середине. Однако, по мере накопления экспериментальных данных и разработке высокоточных методов исследования (рентгеноструктурный анализ) от бутербродной модели отказались. В 1972 г. Сингером и Николсоном предложена новая модель строения биологической мембраны – жидко-мозаичная. Структурную основу мембраны образует двойной слой фосфолипидов, инкрустированных белками. Среди белков различают периферические и интегральные белки. Липиды при физиологических условиях находятся в жидком агрегатном состоянии, что позволяет сравнивать мембрану с фосфолипидным морем, по которому плавают айсберги белков. Кроме фосфолипидов и белков в биологичекой мембране содержаться и другие химические соединения: много холестерина (много в биологических мембранах животных клеток), гликолипиды и гликопротеины. 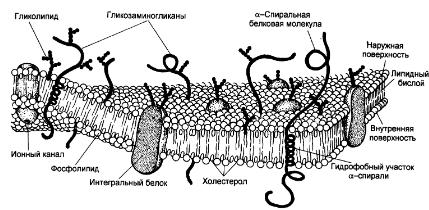 Жидко-мозаичная модель строения биологических мембран в настоящее время общепринята, но как всякая модель она дает упрощенную картину строения мембран. В частности, обнаружено, что белковые молекулы не всегда свободно плавают в липидном море и могут быть заякорены на внутриплазматические структуры клетки, которые составляют ее скелет (цитоскелет). Эти структуры образованы двумя элементами: микрофиламентами и микротрубочками. Микрофиламенты – тонкие нити из актиноподобного белка. Актин – основной белок тонких мышечных волокон, которые пронизывают всю цитоплазму клетки. Микротрубочки – полые цилиндры, d=300 нм., состоят из особенного белка тубулина, основная функция цитоскелета – опорная, т.е. поддержание жесткой конструкции, но, например, в нервных клетках микротрубочки составляют часть системы быстрого механизма переноса веществ. Кроме того, установлено, что не все липиды в мембране расположены по принципу бислоя. Так, показано, что в липидной фазе содержаться участки, где липидные молекулы не образуют двойной слой. Полисахариды связаны с молекулами периферических белков, располагаются на наружной поверхности мембраны, также с интегральными белками. 83. Виды пассивного транспорта веществ в биологических мембранах. Различают несколько видов пассивного переноса веществ через мембрану: -простая диффузия; -перенос через поры (каналы) в мембране; -облегченная диффузия (транспорт веществ с помощью переносчиков, которыми являются особые белки). Пассивный транспорт – это перенос веществ из мест с большим значением его электро-химического потенциала в места с меньшим значением. Идет с уменьшением энергии Гиббса, поэтому этот процесс может осуществляться самопроизвольно без затрат внешней энергии. 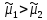 , ,  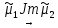 На движение ионов с зарядом z оказывают влияния электричесие поля с напряженность 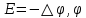 -потенциал. -потенциал.Интенсивность потоков ионов будет зависеть от подвижности ионов данного сорта U. Самоподвижность характеризует скорость движения ионов под действием электрического поля. Подвижность ионов и коэффициент диффузии D(1) связаны через уравнение Эйнштейна (2) следующим образом: 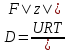 , z-заряд по абсолютной величине. , z-заряд по абсолютной величине.Полная плотность потока ионов данного сорта, которая учитывает наличие диффузионных сил, описывается в уравнении (1) и сил электрического поля: 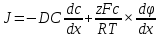 (4). (4).Отсюда с учетом (2), при котором D=URT, полный поток ионов определенного сорта: 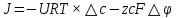 (5). (5).Это уравнение называют электро-диффузионное уравнение Нернста-Планка и показывает, что могут быть две причины переноса ионов при пассивном транспорте: 1 причина – градиент концентрации 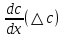 ; ;2 причина – градиент электрического потенциала 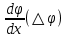 ; ;Знак «-» показывает, что градиент концентрации вызывает перенос вещества от мест с большей концентрацией, а градиент электрического потенциала вызывает перенос «+» зарядов от мест с большим потенциалом к местам с меньшим потенциалом. Но в отдельных случаях в следствие сопряжения 2х причин может происходить пассивный перенос веществ от мест с меньшей концентрацией к местам с большей концентрацией, если второй член уравнения будет больше первого по модулю. Может происходить перенос веществ от мест с меньшим потенциалом к большим, если первый член уравнения по модулю больше второго. Основные разновидности простой диффузии Простая диффузия – самопроизвольный процесс перемещения вещества по градиенту концентрации. Диффузия вещества через липидный бислой вызывается градиентом концентраций в мембране. 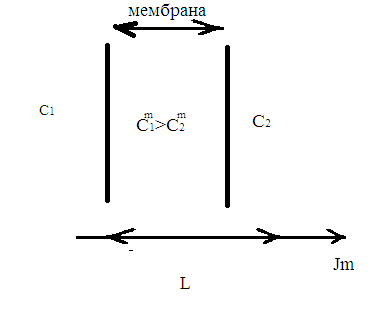 C1- концентрация у одной из сторон мембраны вне ее; C2- концентрация у другой поверхности мембраны вне ее; C1m- концентрация у одной из сторон мембраны, но внутри нее; C2m- концентрация у другой поверхности мембраны, но внутри нее; L- толщина мембраны Плотность потока вещества по закону Фика: 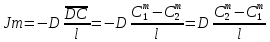 (6) (6)Т.к. измерить концентрации 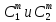 трудно, на практике используют формулу, которой связывают плотность потока вещества через мембрану с концентрацией этого вещества не внутри мембраны, а снаружи в растворах около внешней поверхности мембраны (C1 и C2), тогда формула плотности потока вещества: трудно, на практике используют формулу, которой связывают плотность потока вещества через мембрану с концентрацией этого вещества не внутри мембраны, а снаружи в растворах около внешней поверхности мембраны (C1 и C2), тогда формула плотности потока вещества: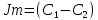 (7); P- коэффициент проницаемости мембраны (7); P- коэффициент проницаемости мембраныКоэффициент проницаемости мембраны (P) зависит от свойств мембран и переносимых через нее веществ; если считать концентрацию вещества у поверхности в мембране прямо пропорциональной концентрациям у поверхности вне нее, то 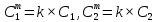 , k – коэффициент пропорциональности. , k – коэффициент пропорциональности.Величина k показывает какую часть концентрации у поверхности вне мембраны составляет концентрация у поверхности, но внутри нее. Подставляем в уравнение Фика (6) значения 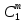 и и 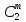 , получим выражение плотности потока: , получим выражение плотности потока: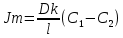 (8) (8)Из уравнений (7) и (8) находим, что коэффициент проницаемости 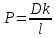 . .Он тем больше, чем больше коэффициент диффузии (D), т.е. чем меньше вязкость мембраны. P больше, чем тоньше мембрана ( Хорошо растворим в фосфолипидном бислое неполярные вещества (органические жирные кислоты, эфиры), поэтому они легко проникают через липидный бислой. Плохо проходят полярные водорастворимые вещества, соли, основания, сахара, аминокислоты, спирты. Другой разновидностью простой диффузии является диффузия через липидные/белковые каналы в мембране. Чрез них проникают молекул, нерастворимые в липидах вещества и водорастворимые гидратированные ионы (окружены водой). Для жирно растворимых веществ и ионов мембраны – молекулярное сито, т.е. чем больше размер молекулы, тем меньше проницаемость мембраны для этого вещества. Облегченная диффузия Подвижные переносчики и каналообразующие объекты, происходит с участием особых белков-переносчиков. Их функциональное назначение – обеспечение подвижности молекул через липидный бислой мембраны. Соединяясь с транспортируемым веществом, которое само проходить через мембрану не может, переносчик перетаскивает его сквозь бислой. Присутствуют в биологической мембране переносчики – высокомолекулярные белки, которые встроены в липидный бислой и ориентированы в нем определенным образом. С помощью переносчика, транспортирующего аминокислоты и органические кислоты, моносахариды, нуклеотиды наиболее изучены переносчики мембран эритроцитов человека. Их мембраны содержат набор разнообразных специализированных транспортных систем, относительное содержание которых невелико. Пример 1: переносчик глюкозы. Количественное содержание переносчика глюкозы в 1 клетке = 2% от всех мембранных белков эритроцитов, но эффективность действия этой системы высоко, т.к. гидрофобный липидный бислой почти непроницаем для гидрофильной молекулы глюкозы, скорость ее прохождения через искусственный липидный бислой незначительный. Переносчик глюкозы увеличивает скорость этого процесса в живой мембране от 100 тысяч до 1 млн. раз. Переносчик глюкозы – интегральный белок, пронизывающий биологическую мембрану эритроцита, образуется пора. Селективные центры связывания глюкозы расположены внутри этой поры, они одинаково доступно как с внутренней, так и с внешней стороны биологической мембраны. 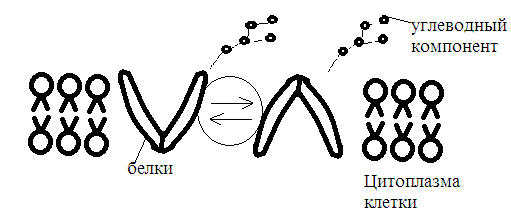 Механизм действия этого переносчика основан на способности белкового агрегата находиться в двух структурных состояниях. В первом он связывает глюкозу, а во втором высвобождает ее с противоположной стороны мембраны. Пример 2-я группа переносчиков Проникновение многоклеточных соединений через мембрану может осуществляться мембранно-активными веществами, которые способны переносить ионы щелочных и щелочноземельных металлов – ионофоры. Термин объединяет мембранно-активные вещества гидрофобной природы, которые способствуют переносу ионов через липидные барьеры в биологической мембране. К ним относят макроциклические соединения с большим количеством кислорода. Все они отличаются способностью избирательно связывать ионы металла и образуют с ними липидно-растворимые комплексы. Эти комплексы ионофоров с металлом образуются при взаимодействии с атомами O2, которые равномерно встроены в циклический скелет молекул и ориентирован соответствующим образом. В зависимости от механизмов взаимодействия ионофоров с биологической мембраной они бывают: -подвижные вещества переносчики; -вещества канало-формеры. Подвижные вещества переносчики. Классический пример подвижного переносчика – валиномицин – антибиотик, который продуцируется различными штаммами гриба Streptomyces. Специфика его биологической активности состоит в том, что он необычайно высоко избирателен по отношению к ионам K. Он обладает уникальной способностью узнавать K среди различных катионов, отличать его в том числе от Na. Избиратльность такого комплексообразователя высока и составляет примерно 10000, если ее оценивать по отношению проводимости через биологическую мембрану по K и по Na. Причина такого действия состоит в особенной пространственной структуре молекул валиномицина, а именно его цикличность. Молекул построена из 3-х фрагментов, содержащих регулярно чередующиеся остатки аминокислот и оксикислот (D- и L- валина и L-молочной и D-изовалериановой кислот). Пространственная организация валиномицина определяется образованием внутримолекулярных H-связей между атомами O2 и карбоксильной группой; и атомами H NH-групп. Это создает упорядоченную пространственную структуру в форме манжетки, которая имеет размер. 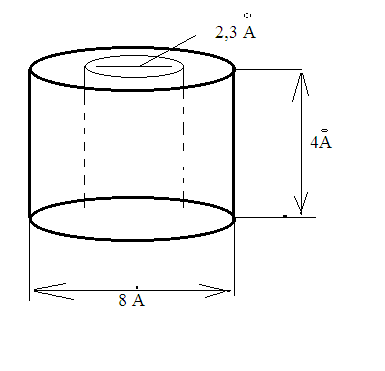 Свободные карбонильные группы направлены к внешним сторонам такого кольца (все наружу). При появлении катиона K эти карбонильные группы приобретают ориентацию наоборот (все внутрь). В итогу, внутри этого браслета полость = 2,3 Å. Наружу теперь ориентированы углеводородные остатки органических кислот (молочной и изовалериановой). Такая структура имеет липофильный характер молекулярной поверхности валиномицина и делаем образовавшийся комплекс (валиномицина+K) легко растворимым в липидном бислое мембраны. Образовавшееся пространство внутренней полости молекулы валиномицина идеально соответствует только размеру одного катиона K (его ионный радиус 1, 33Å). Это обеспечивает избирательное его связывание. Ионный радиус Na=0,95 Å. Внутри молекулы H-связи валиномицина придают жесткость этой полости в браслете, специально приспособленных только для ионов K. Эту полость трудно сжать до размера Na т.к. взаимодействие всех 6 атомов O2 c катионом Na энергетически невыгодно. Это причина высокой избирательности валиномицина по отношению к иону K. Пассивный транспорт ионов K с помощью валиномицина происходит с большой скоростью. Так, одна молекула валиномицина переносит через биологическую мембрану 10 тыс ионов K в секунду. Молекула валиномицина оказывается у поверхности мембраны, захватывая K из окружающей среды, в результате чего он диффундирует в мембране. Если концентрация K по обе стороны мембраны одинакова, то переноса не происходит. 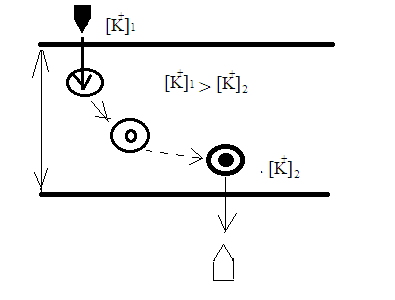 Если с одной из сторон концентрация K снаружи больше K с внутренней сторону, то осуществляется перенос и наоборот. Т.о. происходит диффузия из мест с большей концентрацией в места с меньшей концентрацией. Вещества канало-формеры антибиотики группы грамицидин – пептид, состоящий из 15 остатков гидрофобных аминокислот. Грамицидин – смесь антибиотиков, которые содержат 85% грамицидина А. Пространственная структура грамицидина – β-спиральная конформация полипептидной цепи. Длина такой спирали ½ толщины мембраны. Ее структура создает возможность для образования димеров при объединении их N-концов. 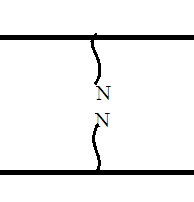 В этой структуре гидрофобные боковые цепи расположены на внешней поверхности такого спирализированного димера образуют липофильную молекулярную поверхность, которая способна к контакту с углеводородным радикалом липидов. Т.о. формируется трансмембранный ионный канал, внутри которого обращены полярные группы и образуется пора с d=5 Å. Ее функция – избирательный канал для одновалентных электронов. Время жизни единичного канала, который обеспечивает перенос ионов в открытое состояние, примерно секунда. За это время он обеспечивает перемещение более 10 млн одновалентных катионов. Передача иона может осуществляться от одной молекулы переносчика к другой по эстафете, поэтому перенос еще называют эстафетным. Отличие облегченной диффузии от простой: Перенос веществ с участием переносчика происходит значительной быстрее Облегченная диффузии обладает свойством насыщенности, т.е. при повышение концентрации с одной стороны плотность потока вещества возрастает лишь до некоторого значения, когда все молекулы переносчики будут заняты. 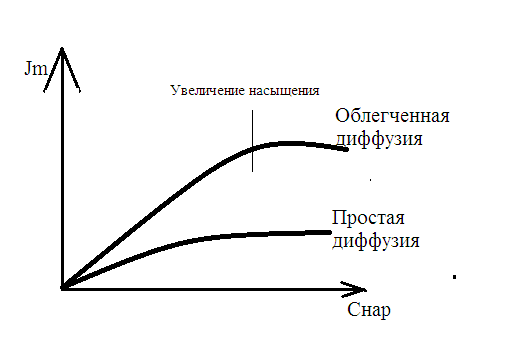 При облегченной диффузии наблюдается конкуренция веществ переносчиков к транспортируемым веществам (когда переносчик переносит разные вещества) и строгая избирательность. Процессы облегченной диффузии чувствительны к действию определенных веществ – ингибиторов, которые блокируют транспорт через биологическую мембрану. Если транспорт обладает перечисленными особенностями, то имеет место облегченная диффузия. |