|
|
ответы экзамен. Эволюция пищеварительной системы и особенности питания у представителей разных классов позвоночных животных
2.3. Са2+-АТФаза
Кальциевые АТФазы, входящие в состав цитоплазматических мембран и внутриклеточных мембран, различаются по ряду свойств. Все Са-АТФазы представляют собой мономерные белки, то есть состоят из единственной полипептидной цепи, но несколько различаются по молекулярной массе. Так, Са-АТФаза саркоплазматического ретикулума имеет молекулярную массу 108 кД, а плазматическая Са-АТФаза – 120 кД. Лучше всего изучена Са-АТФаза саркоплазматического ретикулума поперечнополосатых мышц.
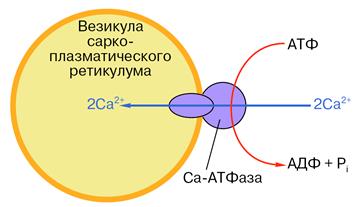
Рис. 13. Схематическое изображение везикулы саркоплазматического ретикулума со встроенной молекулой Са-АТФазы. Во внешнюю среду (цитоплазму) обращена головка фермента диаметром около 9 нм. С ней связываются АТФ и ионы кальция. Мембрану пронизывает канал, по которому, как полагают, кальций переносится при гидролизе АТФ. Более подробная схема строения фермента показана на рис. 15.
Механизм переноса ионов кальция.Каждый цикл переноса включает в себя как минимум три стадии:
1) захватит частицы с одной стороны мембраны,
2) перенес ее через мембрану (транслоцирование),
3) выпуск частицы с другой стороны.
Осуществление этих стадий сопряжено с расходом энергии, и, следовательно, одновременно что-то должно происходить с АТФ. Сама молекула АТФ тоже должна быть захвачена (1) и гидролизована с запасанием энергии и расходом ее на перенос кальция (2), а продукты (АДФ и фосфат) должны перейти из связанного с ферментом состояния в водный раствор (3). В каждом цикле фермент одновременно использует не один, а два субстрата (внутриклеточный кальций и АТФ) и образует три продукта: кальций, накопленный внутри везикул эндоплазматического ретикулума, АДФ и ортофосфат. Усилиями многих ученых была в общих чертах расшифрована последовательность стадий при работе Са-АТФазы (рис. 14), которая включает в себя все перечисленные выше этапы. Работа насоса замечательна еще и тем, что стадии переработки АТФ как бы чередуются со стадиями переноса Са2+. Эти стадии перечислены ниже:
1. связывание двух ионов кальция на поверхности АТФазы, обращенной в цитоплазму (или наружу в изолированных пузырьках СР);
2. связывание на той же поверхности молекулы АТФ;
3. фосфорилирование белка (образование фосфофермента) и высвобождение АДФ;
4. высвобождение ионов кальция с поверхности АТФазы, обращенной внутрь пузырьков СР; связывание магния;
5. гидролиз фосфатной связи и отщепление ионов магния;
6. переход молекулы фермента в исходное состояние (центры связывания кальция оказываются опять на поверхности пузырьков СР).
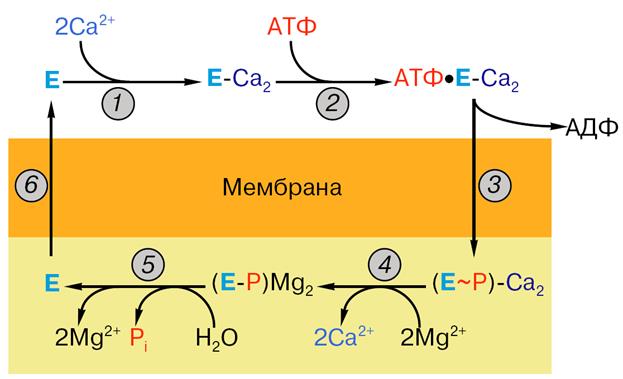
Рис. 14. Последовательность стадий работы Са-АТФазы: 1 – связывание ионов кальция, 2 – связывание АТФ, 3 – образование фосфо-фермента, 4 – отщепление ионов кальция, 5 – гидролиз фосфо-фермента, 6 – возвращение фермента в исходное состояние. Остальные объяснения даны в тексте.
Связывание ионов кальция (1 стадия). К активному центру белка Са2+-АТФазы присоединяется 2 иона Са2+. Установлено, что каждый шестой центр связывания на поверхности молекул АТФазы занят ионами кальция при той чрезвычайно низкой (100 нМ) их концентрации, которая типична для внутриклеточного содержимого.
Связывание АТФ (2 стадия). На поверхности АТФазы имеются центры связывания для двух ионов кальция и одной молекулы АТФ, обладающие высоким сродством к субстрату. Они взаимодействуют между собой, так как связывание Са2+ запускает гидролиз АТФ. При этом было показано, что гидролиз АТФ начинается только после того, как оба иона кальция присоединятся к своим участкам связывания.
Фосфорилирование белка (стадия 3).Гидролиз АТФ осуществляется Са-АТФазой в три этапа. Вначале происходит связывание АТФ, затем фосфорилирование белка и отщепление АДФ и, наконец, гидролитическое расщепление белок-фосфатной связи и высвобождение ортофосфата.
Как известно, при гидролизе АТФ высвобождается большое количество энергии, благодаря чему связь между фосфатом и АДФ в молекуле АТФ называют макроэргической (богатой энергией). Обратимость процесса фосфорилирования белка означает, что и связь фосфата с аспарагиновым остатком в фосфорилированном белке (на рис. 14 она обозначена знаком
) тоже богата энергией, которая высвобождается при ее гидролизе. Именно эта энергия и тратится на активный перенос ионов кальция.
Высвобождение ионов кальция (стадия 4).Высокоэнергетическая (способная передавать остаток фосфорной кислоты на АДФ) форма фосфорилированной АТФазы стабильна только в присутствии миллимолярных (то есть сравнительно высоких) концентраций ионов Са2+. При меньших концентрациях Са2+ происходит вытеснение ионов Са2+ из Са-связывающих центров фосфофермента ионами Mg2+ (которые присутствуют в среде и без которых Са-АТФаза не работает), ионы кальция при этом выходят в окружающий раствор. Эта стадия работы АТФазы (гидролиз ЕР) – важнейшая в цикле переноса ионов кальция и заслуживает пристального рассмотрения.
Вытеснение ионов Ca2+ из Са-связывающих центров высокоэнергетического фосфопроизводного белка ионами Mg2+ происходит, как выяснилось, не одномоментно, а в два этапа: сначала отщепляется кальций и только потом происходит гидролиз фосфатной связи (отщепление неорганического фосфата):
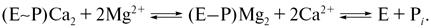
Перенос кальция через мембрану (транслокация).В везикулах саркоплазматического ретикулума молекулы Са-АТФазы ориентированы строго определенным образом, так что связывание ионов кальция и АТФ происходит с наружной стороны пузырьков, а высвобождение кальция – с внутренней. Изучение связывания ионов кальция на разных стадиях работы Са-АТФазы показало, что в нефосфорилированном состоянии Са-связывающие центры АТФазы доступны для ионов Са2+ только с внешней стороны пузырьков и недоступны с внутренней. После фосфорилирования фермента Са-связывающие центры становятся доступными с внутренней стороны и недоступными с внешней. Таким образом, фосфорилирование приводит к переносу центров связывания кальция через мембрану (транслокации). Поскольку перенос ионов осуществляет белковая молекула, очевидно, какие-то ее части должны перемещаться или, как принято говорить, должно происходить изменение конформации белковой молекулы. Одновременно происходит изменение сродства центров связывания к ионам кальция.
Завершение цикла – гидролиз фосфофермента (стадии 5 и 6).Магниевый комплекс энзимфосфата быстро гидролизуется, и фермент приобретает свои исходные свойства (иными словами, восстанавливается исходная конформация ферментного белка). При этом на поверхности фермента, обращенной наружу, вновь появляются центры связывания кальция с высоким сродством. Очевидно, что гидролиз Е-Р приводит, во-первых, к освобождению центров связывания от магния, а во-вторых, к их обратной транслокации (стадия 6). При этом на поверхности фермента, обращенной наружу, центры связывания кальция приобретают вновь высокое сродство к этим ионам. Таким образом, дефосфорилирование Е-Р приводит к пространственным перемещениям участка белковой молекулы и изменениям энергии связывания ионов точно так же, как и фосфорилирование АТФазы, но в противоположном направлении. Цикл работы фермента замыкается (см. рис. 14).
Строение Са2+-АТФазы.Многие ферменты получены в виде кристаллов, и на основании рентгеноструктурного анализа воссоздана их подробная пространственная структура, а подчас и структура их комплексов с субстратами и ингибиторами. К сожалению, транспортные АТФазы, нерастворимые в воде и работающие в составе мембран, не удается получить в виде настоящих кристаллов. Тем не менее, многое об их структуре все же известно, включая последовательность аминокислот в полипептидной цепи, локализацию мест связывания ионов и АТФ в полипептидной цепи и расположение определенных участков цепи по отношению к мембране.
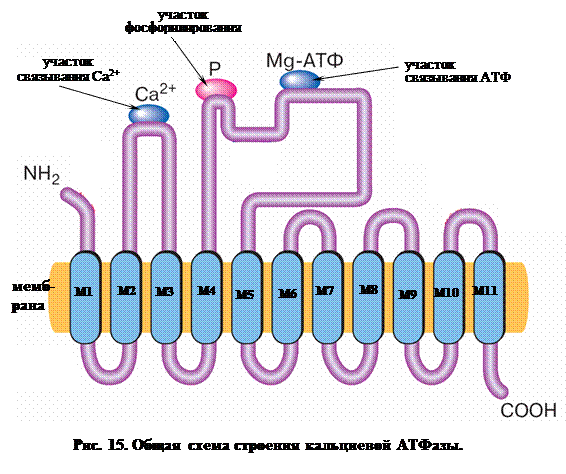
На рис. 15 приведено схематическое изображение Са-АТФазы саркоплазматического ретикулума скелетных мышц. Фермент пронизывает мембрану 11-ю α-спиральными участками, большая часть которых соединена снаружи короткими полипептидными связками, за исключением двух протяженных гидрофильных (то есть хорошо растворимых в воде) петель на стороне цитоплазмы. Более короткая петля расположена между α-спиралями М2 и МЗ, более длинная – между α-спиралями М4 и М5. Длинная петля содержит АТФ-связывающий участок, включающий остаток аспарагиновой кислоты, к которому присоединяется фосфат. Связывание ионов Са2+ происходит на участке, образованном малой петлей (между α-спиралями М2 и МЗ), возможно с участием аминокислотных остатков, прилежащих к спиралям Ml и М4. В местах связывания собрано несколько остатков аспарагиновой кислоты, несущих отрицательные заряды.
86. Биофизический механизм восприятия звуковых колебаний в отделах органа слуха человека.
Наружное ухо состоит из ушной раковины, наружного слухового прохода, которое имеет длину 2,7 см, наполняющий органную трубку, которая закрыта с одного конца барабанной перепонкой. Резонанс наблюдается, если длина резонатора (слухового прохода) имеет 1/4 длины волны. Наружной слух проход поэтому имеет резонансную частоту около 3.000 Гц, что соответствует максимальной чувствительности слуха человека.
Среднее ухо - является устройством, которое трансформирует звуковые колебания воздуха в звуковые колебания жидкой среды внутри уха, то есть среды которая имеет больше энергию, чем больше воздух. Чтобы привести в движение эту жидкость нужно получить выигрыш в давлении. Основной элемент среднего уха – слуховые косточки.
Выигрыш в резонаторе достигается благодаря:
система слуховых косточек – молоточек, наковальня, стремечко работает как рычаг, который обеспечивает выигрыш в силе в 1,3 раза
площадь барабанной перепонки у человека (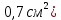 значительно больше площади овального окна внутри уха( значительно больше площади овального окна внутри уха(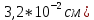 , то есть меньше в 20 раз, поэтому объём мембраны и связь их слуховых косточек выполняет функцию трансформатора резонатора. , то есть меньше в 20 раз, поэтому объём мембраны и связь их слуховых косточек выполняет функцию трансформатора резонатора.
Другая важная функция среднего уха является защита внутреннего уха от чрезмерной механических нагрузки при воздействии очень громкого звука. Это достигается за счет изменения характера колебания стремячка в зависимости от силы звука.
При пороговым значении (минимамума) звука резонатор стремячка колеблется, как поршень. Абсолютное смещение стремечка очень малы и они повторяют движения барабанной перепонки. Смещение барабанной перепонки при пороговом значении Резонатора является 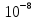 миллиметра, это меньше чем радиус атома. Если сила звука увеличить до 150 дБ, то появляется ощущение боли, а при больше 150 дБ барабанная перепонка разрушаются, хотя её смещение достигает всего 1 мм. При возрастании силы звука характер колебания стремячка меняется. Так как при средней силе звука стремечко начинает совершать колебательные движения вокруг вертикальной оси у одного конца овального окна. миллиметра, это меньше чем радиус атома. Если сила звука увеличить до 150 дБ, то появляется ощущение боли, а при больше 150 дБ барабанная перепонка разрушаются, хотя её смещение достигает всего 1 мм. При возрастании силы звука характер колебания стремячка меняется. Так как при средней силе звука стремечко начинает совершать колебательные движения вокруг вертикальной оси у одного конца овального окна.
При очень громком звуке стремечко начинает совершать вращательные движения вокруг горизонтальной оси овального окна так, когда один конец стремячка вдавливается в овальное окно, а другой движется в противоположную сторону.
Таким образом предотвращается избыточно сильное движение жидкости во внутреннем ухе. Такое изменение движения стремечка имеет решающее значение для защиты внутреннего уха от повреждений при внезапных резких звуках, когда не успевает осуществится любой рефлекторный механизм защиты.
Внутреннее ухо в отличие от наружного и среднего заполнено жидкостью. Для слуха важна только улитковая часть, она имеет форму спирали и образуют 2,5 витка.
Длина развернутой улитки около 35 мм, а ее объем — около 100 мкл (т. е. равен объему двух капель воды). В улитке располагаются три параллельных наполненных жидкостью канала. Вестибулярный и барабанный каналы заполнены перилимфой и соединены в вершине улитки маленьким отверстием – геликотремой.
Эти два канала отделены друг от друга не сообщающимся с ними улитковым каналом, заполненным эндолимфой и отделенным от вестибулярного канала очень тонкой мембраной Рейснера, а от барабанного канала — базилярной мембраной. К базилярной мембране примыкает кортиев орган, содержащий рецепторные клетки и нервные окончания. Рецепторами звука считаются волосковые клетки. Характер колебаний базилярной мембраны зависит от частоты. При очень низких частотах волны давления, передаваемые стремечком от барабанной перепонки, заставляют перилимфу двигаться вперед и назад через геликотрему, вызывая колебания круглого окна. Такие низкочастотные колебания почти не приводят в движение базилярную мембрану. При более высоких частотах, например 30 Гц, волны давления из-за инерционности жидкости стремятся распространиться прямо через базилярную мембрану, приводя ее в движение. Базилярная мембрана очень неоднородна по длине, так от овального окна к вершине улитки она уширяется и утолщается. Рядом со стремечком она уже, легче и имеет примерно в 100 раз большее значение модуля упругости, чем у вершины. Благодаря неоднородным механическим свойствам базилярной мембраны волны разной частоты приводят в движение различные ее участки. Низкие частоты (менее 100 Гц) вызывают колебания наиболее массивной части мембраны около геликотремы. Высокие частоты (8000 и более Гц), наоборот, приводят в движение участок мембраны вблизи овального окна. Для частоты 1600 Гц максимум колебаний лежит около середины улитки.
Преобразование механических колебаний в нервном импульсе.
Колебания базилярной мембраны вызывает деформацию волосковых клеток, располагающихся в кортиевом органе внутри улиткового канала. Волосковые клетки цилиндрической формы, сужены на одном конце. Суженая часть имеет 2 типа волосков:
одиночная длинная неподвижная ресничка – киноцилия
стереоцилия
От 40 до 60 стереоцилий объединены в пучки, их кончики соединяются с киноцилией тонкими полипептидными нитями. Волосковые клетки чрезвычайно чувствительны к механическим стимулам. Деформация волосковых клеток приводит к нарушению ионного равновесия в их цитоплазматических мембранах. Пороговые сдвиги вызывают на мембране волосковых клеток рецепторный потенциал, амплитуда приблизительно 100 микро Вольт. Знак потенциала зависит от направления движения стереоцилий относительно киноцилий. Так если пучок стереоцилий наклонен по направлению к киноцилии наблюдается деполяризация, если в противоположную сторону – гиперполяризация. Возникающие изменения потенциала действуют на немиелинизированные дендриты афферентных нейронов, которые находятся в контакте с волосковыми клетками.
В результате происходит возбуждение слухового нерва, звук определенной частицы приводит в движение и вызывает нервный потенциал от определенной части базилярной мембраны.
В центральной нервной системе имеется механизм, обостряющий ощущение звуковой частоты так, что в диапазоне 60—1000 Гц человеческое ухо может различать частоты, отличающиеся на 2—3 Гц.
Слуховой аппарат человека исключительно чувствителен. Как отмечалось выше, пороговые колебания барабанной перепонки составляют мм, а вызываемые этим колебания базилярной мембраны еще меньше по амплитуде. Поэтому Кортиев орган не имеет кровеносных сосудов, чтобы пульсации кровяного давления не приводили в движение волосковые клетки и не вызвали слуховых ощущений.
В аудиометрии существует понятие пространственная локализация звука – это способность человека устанавливать месторасположение и направление движения источника звука. Иногда это субъективную способность называют бинауральным (двуухим) эффектом. Благодаря разности восприятия звуковых характеристик правого и левого уха и их ориентации на звучащий объект удается определить его пространственное и временные координаты. Для этого анализируются тональные, громкостные и фазовые различия звука между ушами. Для оценки звуковой функции на низких частотах служит разность во времени, а для высоких частот разница в громкости. Минимальная улавливаемая человеком разница во времени прихода звука между ушами составляет ниже 30 мс. При высокочастотных (например 5000 герц) длина волны звука приблизительно 6,8 см, в связи с чем звук не может обогнуть голову и попасть в противоположную ухо. Механизм пространственного распространения звука основан на различиях громкости. Дифференциальный порог громкости:3,1-1дБ. В результате действия 2 механизмов точность обнаружения направления звука, то есть разность между истинным и субъективным радиусом составляет 1 градуса до 3 градусов. Наиболее распространенным методом изучением слуха является аудиометрия. С помощью аудиометра измеряются пороги слышимости при использовании чистых тонов. Полученные величины сравниваются с нормальной величины порога. В диапазоне слышимых частот человека различают направление, в котором происходит изменение частоты, если это изменение достаточно велико. Наибольшая частота звука, которую слышат люди различна и изменяется более трех раз (от 8000 до 2.500 Гц).
Нормальным считаются слух при величине дифференциального порога силы звука (от 0,8 до 2 дБ), величина меньше 0,8дБ служит признаком расстройства слухового анализатора в виде повышения его чувствительности к нарастанию громких звуков. Величина ниже 0,8 дБ служат отборочным тестом для выявления музыкально одаренных детей. По мере старения у человека порог восприятия чистых тонов повышается. В результате длительного воздействия громких шумов для людей всех возрастов эти пороги повышаются. Увеличение пороговой слуховой чувствительности называется - тугоухостью. Нарушением слуха страдают приблизительно 3%. В зависимости от анатомо-функциональной обусловленности и этиогенеза в виды тугоухости делятся на: кондуктивную (расстройство звукопроведения) и нейросенсорную (расстройство звуковосприятия).
|
|
|
 Скачать 7.56 Mb.
Скачать 7.56 Mb.