ответы экзамен. Эволюция пищеварительной системы и особенности питания у представителей разных классов позвоночных животных
 Скачать 7.56 Mb. Скачать 7.56 Mb.
|

|
Колонизация – процесс размножения микроорганизмов на поверхности эпителия. Для успешной колонизации очага первичного инфицирования бактерии должны выдержать действие многочисленных и разнообразных микробицидных факторов хозяина. Для защиты от них микроорганизмы активно используют ряд структур (капсула, поверхностные протеины) и синтезируемых веществ (экзоферменты). Гидрофильность капсул затрудняет их поглощение фагоцитами, а само капсульное вещество защищает бактерию от воздействия лизосомальных ферментов и токсичных оксидантов, выделяемых фагоцитирующими клетками. Кроме того, бактерии при поглощениии легко «снимают с себя» капсулу и избегают контакта с фагоцитом. Пенетрация – процесс проникновения микроорганизмов внутрь клеток макроорганизма. Это характерно для вирусов, некоторых патогенных бактерий (шигеллы, энтероинвазивные кишечные палочки). Микроорганизмы размножаются внутри клеток, которые гибнут, вызывая нарушение целостности эпителиального покрова (эрозии). Инвазия – способность микроорганизмов проникать через слизистые и соединительнотканные барьеры в подлежащие ткани. Этот процесс обеспечивают жгутики, ферменты. Например, гиалуронидаза (Clostridium perfringens, некоторые Streptococcus sp., некоторые Staphylococcus sp.) расщепляет гиалуроновую кислоту, входящую в состав межклеточного вещества, что повышает проницаемость слизистых оболочек и соединительной ткани. Нейраминидаза (Vibrio cholerae, Yersinia sрp., Pasterella sрp., Streptococcus sрp., некоторые Clostridium sрp.) разрушает гликозидные связи, отщепляя концевые сиаловые кислоты от углеводов. Сиаловые кислоты деполимеризуют поверхностные структуры эпителиальных и других клеток организма, разжижают носовой секрет, слой слизи (муцина) кишечника, способствует распространению не только через слизистую оболочку, но и внутрь клеток. Агрессия – способность патогенных микроорганизмов размножаться в организме хозяина и противостоять его защитным механизмам. Агрессия осуществляется за счет структур клеточной стенки: капсулы, клеточной стенки, липополисахаридов (ЛПС) Грам- бактерий , которые подавляю миграцию лейкоцитов, препятствуют фагоцитозу. Для подавления иммунитета патогенные микроорганизмы продуцируют различные экзоферменты: протеазы – разрушают иммуноглобулины (антитела), плазмокоагулазу – свертывает плазму крови, фибринолизин – растворяющий сгустки фибрина, способствуя гематогенному распространению микробов, лецитиназу – расщепляющую лецитин цитоплазматических мембран эукариотических клеток, уреаза H.pylori нейтрализует кислую среду в желудке. Генетические рекомбинации у бактерий: трансформация, трансдукция, конъюгация. Конъюгация бактерий состоит в переходе генетического материала (ДНК) из клетки-донора («мужской») в клетку-реципиент («женскую») при контакте клеток между собой. Мужская клетка содержит F-фактор, или половой фактор, который контролирует синтез так называемых половых пилей, или F-пилей. Клетки, не содержащие F-фактора, являются женскими; при получении F-фактора они превращаются в «мужские» и сами становятся донорами. F-фактор располагается в цитоплазме в виде кольцевой двунитчатой молекулы ДНК, т. е. является плазмидой. Молекула F-фактора значительно меньше хромосомы и содержит гены, контролирующие процесс конъюгации, в том числе синтез F-пилей. При конъюгации F-пили соединяют «мужскую» и «женскую» клетки, обеспечивая переход ДНК через конъюгационный мостик или F-пили. Клетки, содержащие F-фактор в цитоплазме, обозначаются F+; они передают F-фактор клеткам, обозначаемым F" («женским»), не утрачивая донорской способности, так как оставляют копии F-фактора. Если F-фактор включается в хромосому, то бактерии приобретают способность передавать фрагменты хромосомной ДНК и называются Hfr-клетками (от англ. high frequency of recombination — высокая частота рекомбинаций), т.е. бактериями с высокой частотой рекомбинаций. При конъюгации клеток Hfr и клеток F" хромосома разрывается и передается с определенного участка (начальной точки) в клетку F", продолжая реплицироваться. Перенос всей хромосомы может длиться до 100 мин. Переносимая ДНК взаимодействует с ДНК реципиента — происходит гомологичная рекомбинация. Прерывая процесс конъюгации бактерий, можно определять последовательность расположения генов в хромосоме. Иногда F-фактор может при выходе из хромосомы захватывать небольшую ее часть, образуя так называемый замещенный фактор — F'. При конъюгации происходит только частичный перенос генетического материала, поэтому ее не следует отождествлять полностью с половым процессом у других организмов. Трансдукция — передача ДНК от бактерии-донора к бактерии-реципиенту при участии бактериофага. Различают неспецифическую (общую) трансдукцию, при которой возможен перенос любого фрагмента ДНК донора, и специфическую — перенос определенного фрагмента ДНК донора только в определенные участки ДНК реципиента. Неспецифическая трансдукция обусловлена включением ДНК донора в головку фага дополнительно к геному фага или вместо генома фага (дефектные фаги). Специфическая трансдукция обусловлена замещением некоторых генов фага генами хромосомы клетки-донора. Фаговая ДНК, несущая фрагменты хромосомы клетки-донора, включается в строго определенные участки хромосомы клетки-реципиента. Таким образом, привносятся новые гены и ДНК фага в виде профага репродуцируется вместе с хромосомой, т.е. этот процесс сопровождается лизоге-нией. Если фрагмент ДНК, переносимый фагом, не вступает в рекомбинацию с хромосомой реципиента и не реплицируется, но с него считывается информация о синтезе соответствующего продукта, такая трансдукция называется абортивной. Трансформация заключается в том, что ДНК, выделенная из бактерий в свободной растворимой форме, передается бактерии-реципиенту. При трансформации рекомбинация происходит, если ДНК бактерий родственны друг другу. В этом случае возможен обмен гомологичных участков собственной и проникшей извне ДНК. Впервые явление трансформации описал Ф. Гриффите (1928). Он вводил мышам живой невирулентный бескапсульный R-штамм пневмококка и одновременно убитый вирулентный капсульный S-штамм пневмококка. Из крови погибших мышей был выделен вирулентный пневмококк, имеющий капсулу убитого S-штамма пневмококка. Таким образом, убитый S-штамм пневмококка передал наследственную способность капсулообразования R-штамму пневмококка. О. Эвери, К. Мак-Леод и М. Мак-Карти (1944) доказали, что трансформирующим агентом в этом случае является ДНК. Путем трансформации могут быть перенесены различные признаки: капсулообразование, устойчивость к антибиотикам, синтез ферментов. +Изучение бактериальной трансформации позволило установить роль ДНК как материального субстрата наследственности. При изучении генетической трансформации у бактерий были разработаны методы экстракции и очистки ДНК, биохимические и биофизические методы ее анализа. Взаимодействие вирусов с клетками. Взаимодействие вирусов с клетками может осуществляться: а) с образованием вирусного потомства (продуктивный тип взаимодействия); б) без образования вирусного потомства или с незначительным образованием дочерних вирусов (абортивная инфекция); в) с встраиванием вирусной ДНК (РНК) в геном клетки-хозяина (интегративный тип взаимодействия). Примечание. Интегрированная нуклеиновая кислота в составе клеточного генома называется провирусом. Поскольку вирусные инфекции реализуются посредством продуктивного типа взаимодействия вирусов с клетками, остановимся более подробно на процессе репродукции вирусов. В данном процессе выделяют следующие основные этапы: адсорбция вириона на клетке, проникновение вириона в клетку, депротеинизация вириона и освобождение генома вируса, экспрессия вирусного генома и синтез компонентов вирусов: транскрипция, трансляция, репликация, морфогенез (формирование дочерних вирионов), выход нового поколения вирусов из клетки (рис. 7.17). 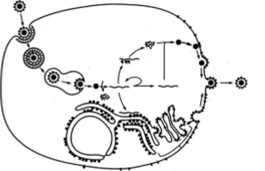 Рис. 7.17. Взаимодействие вируса с клеткой (объяснения см. в тексте) Рассмотрим далее эти этапы. Адсорбция вириона на клетке. Характеризуется высокой специфичностью. Обусловлена взаимодействием белковых рецепторов плазмалеммы чувствительной клетки с прикрепительными белками на поверхности вирусов (у простых вирусов — это белки капсида, у сложных вирусов — эго гликопротеиды суперкаисида). Способность вирусов специфически связываться с определенными клеточными рецепторами называется тропизмом вирусов. Именно этим обстоятельством объясняется избирательное поражение определенными вирусами конкретных биоструктур организма-хозя- ина (клеток, тканей, органов). Например, вирус гепатита В проникает в клетки печени, вирус полиомиелита — в мотонейроны передних рогов спинного мозга. Проникновение вириона в клетку, депротеинизация вириона и освобождение генома вируса. Проникновение вируса может осуществляться двумя способами: путем впячивания плазмалеммы и образования везикулы, содержащей вирус (рецепторзависимый эндоцитоз) и путем слияния плазматической мембраны клетки с сунеркапсидом вириона (при участии специальных белков слияния), в результате внутренняя часть вируса оказывается в цитоплазме клетки. Затем происходят разрушение капсида (депротеинизация) и освобождение вирусной нуклеиновой кислоты. Экспрессия вирусного генома и синтез компонентов вирусов. У ДНК-геномных вирусов включает в себя следующие процессы: вирусная ДНК — транскрипция — трансляция. У РНК-геномных вирусов: вирусная РНК — трансляция. У РНК-геномных ретровирусов: синтез комплементарной ДНК на матрице вирусной РНК — транскрипция — трансляция. Репликация вирусного генома. Сущность: синтез на матрице исходной геномной нуклеиновой кислоты множества копий — будущих вирусных геномов. У большинства вирусов репликация происходит в ядре клетки-хозяина, у некоторых — в цитоплазме. У ДНК-геномных вирусов при участии клеточной ДНК-поли- меразы осуществляется биосинтез множества ДНК-копий (пример: вирус герпеса). У некоторых ДНК-геномных вирусов на матрице ДНК при помощи вирусной ДНК-зависимой РНК-полимеразы синтезируется комплементарная РНК-копия (прегеномная РНК). Затем на пре- геномной РНК при участии обратной транскриптазы (РНК-зави- симой ДНК-полимеразы) собирается комплементарная нить ДНК. Последняя с помощью ДНК-полимеразы достраивается до двухцепочечной молекулы. Таким образом происходит тиражирование копий двунитевых ДНК-геномов вирусов (пример: вирус гепатита В). У РНК-геномных вирусов с помощью вирусной РНК-зависи- мой РНК-полимеразы на матрице геномной РНК синтезируется комплементарная РНК-копия, которая в свою очередь является матрицей для сборки молекул вирусного генома (процесс катализируется тем же ферментом) (пример: вирус гриппа). У РНК-ретровирусов при участии вирусной обратной транскриптазы (РНК-зависимой ДНК-полимеразы) на геномной РНК синтезируется комплементарная ДНК-цепь, которая достраивается до двухцепочечной молекулы с помощью ДНК-полимеразы. Такая ДНК-коиия вирусного генома интегрируется в ДНК клетки- хозяина, где с нее во множестве копий синтезируется геномная РНК вируса (реакция катализируется ДНК-зависимой РНК-поли- меразой). Примечательно, что обратная транскриптаза отличается невысокой специфичностью при сборке цепи ДНК на матрице РНК. Этим объясняется выраженная изменчивость некоторых вирусов, в частности вируса иммунодефицита человека. В результате вышеперечисленных процессов (транскрипция, трансляция, репликация вирусного генома) образуются все компоненты вирусного потомства — геном, геномные белки, белки капсида, а также матриксный белок и гликопротеиды (для сложных вирусов). Процесс формирования дочерних вирионов осуществляется путем самосборки и называется морфогенезом. Выход нового поколения вирусов из клетки может происходить двумя способами. 1. При инфицировании клетки простыми вирусами вирусное потомство покидает клетку после ее лизиса. 2. При инфицировании клетки сложными вирусами вирусные частицы-предшественники (нуклеиновая кислота + капсид) окружаются модифицированной плазматической мембраной клетки, в которую встроены вирусные белки-гликопротеиды, и отпочковываются. При этом клетка-хозяин, как правило, сразу не погибает, а продолжает выделять новые поколения вирусов до истощения ее пластических и энергетических ресурсов. БИР Особенности первых делений зиготы. Сравнение жизненных циклов соматических клеток и бластомеров После сближения женского и мужского пронуклеусов, которое продолжается у млекопитающих около 12 ч, образуется зигота — одноклеточный зародыш. Уже на стадии зиготы выявляются презумптивные зоны (лат. presumptio — вероятность, предположение) как источники развития соответствующих участков бластулы, из которых в дальнейшем формируются зародышевые листки. Дробление и образование бластулы Дробление — последовательное митотическое деление зиготы на клетки (бластомеры) без роста дочерних клеток до размеров материнской. Образующиеся бластомеры остаются объединенными в единый организм зародыша. В зиготе образуется митотическое веретено между отдаляющимися к полюсам центриолями, внесенными сперматозоидом. Пронуклеусы вступают в стадию профазы с формированием объединенного диплоидного набора (Метод выявления презумптивных зон предложен немецким эмбриологом Фогтом) хромосом яйцеклетки и сперматозоида. Пройдя все остальные фазы митотического деления, зигота разделяется на две дочерние клетки — бластомеры. Вследствие фактического отсутствия G1-периода, во время которого происходит рост клеток, образовавшихся в результате деления, клетки гораздо меньше материнской, поэтому и величина зародыша в целом в этот период независимо от числа составляющих его клеток не превышает величину исходной клетки — зиготы. Все это позволило назвать описываемый процесс дроблением (т.е. измельчением), а клетки, образующиеся в процессе дробления, — бластомерами. Дробление зиготы человека начинается к концу первых суток и характеризуется как полное неравномерное асинхронное. В течение первых суток оно происходит медленно. Первое дробление (деление) зиготы завершается через 30 ч, в результате образуется 2 бластомера, по- крытых оболочкой оплодотворения. За стадией двух бластомеров следует стадия трех бластомеров. +С первых же дроблений зиготы формируются два вида бластомеров — «темные» и «светлые». «Светлые», более мелкие, бластомеры дробятся быстрее и располагаются одним слоем вокруг крупных «темных», которые оказываются в середине зародыша. Из поверхностных «светлых» бластомеров в дальнейшем возникает трофобласт, связывающий зародыш с материнским организмом и обеспечивающий его питание. Внутренние, «темные», бластомеры формируют эмбриобласт, из которого образуются тело зародыша и некоторые внезародышевые органы (амнион, желточный мешок, аллантоис). Начиная с трех суток, дробление идет быстрее, и на 4-е сутки зародыш состоит из 7—12 бластомеров. Уже через 50—60 ч образуется плотное скопление клеток — морула, а на 3—4-е сутки начинается формирование бластоцисты — полого пузырька, заполненного жидкостью Бластоциста в течение 3 сут перемещается по яйцеводу к матке и через 4 сут. попадает в матку. Бластоциста находится в полости матки в свободном виде в течение 2 дней (5-е и б-е сутки), и эта стадия обозначается как свободная бластоциста. К этому времени бластоциста увеличивается благодаря росту числа бластомеров — клеток эмбриобласта и трофобласта — до 100 и более вследствие усиленного всасывания трофобластом секрета маточных желез, а также вследствие активной выработки жидкости самим трофобластом (рис). Эмбриобласт располагается в виде узелка зародышевых клеток («зародышевый узелок»), который прикрепляется изнутри к трофобласту на одном из полюсов бластоцисты и начинается имплантация. В большинстве случаев клеточной пролиферации в период между митозами происходит рост клеток. Клетка увеличивается в объеме почти вдвое и затем делится. Такой рост приводит к увеличению общего объема клеток при сохранении относительно постоянного отношения объема ядра к объему цитоплазмы. В период дробления зиготы объем цитоплазмы не возрастает: огромная масса цитоплазмы зиготы разделяется на все более мелкие клетки. Это деление цитоплазмы яйца, не сопровождающееся ростом, осуществляется путем выпадения G1-периода в интерфазе, тогда как митозы следуют друг за другом с большой скоростью. Темпы увеличения числа клеток в период дробления намного выше, чем на стадии гаструляции. Одним из последствий высокой интенсивности делений в процессе дробления является постепенное уменьшение отношения объема цитоплазмы к объему ядра. Изменение скорости, с которой происходит снижение отношения объема цитоплазмы к объему ядра, у многих типов зародышей является решающим фактором, определяющим время активации некоторых генов. От митотического деления соматических клеток дробление отличается тем, что полученные в результате дробления клетки не растут, а поэтому с каждым следующим делением становятся всё более мелкими, при этом увеличивается только их количество, а зародыш в целом не растет. Получающиеся при дроблении клетки мало дифференцированы и сравнительно однородны. Период синхронных делений дробления характеризуется укороченными клеточными циклами, из которых фактически выпадает пре-синтетический, или G1-период, а также постсинтетический, или G2- период . Глубокие изменения в клеточном метаболизме частично связаны с циклическим чередованием окисленной и восстановленной конформации белков, обусловленной S-S и S-H группами. Свободные сульфгидрильные группы в наибольшем числе имеются в делящихся бластомерах и в наименьшем в то время, когда клетки не делятся. |