ответы экзамен. Эволюция пищеварительной системы и особенности питания у представителей разных классов позвоночных животных
 Скачать 7.56 Mb. Скачать 7.56 Mb.
|

|
37.Отдел Бурые водоросли. Классификация. Жизненный цикл фукуса. Отдел бурые водоросли (Phaeophyta) насчитывает около 1500 видов. Бурые водоросли обитают практически только в морях (в пресных водоемах встречается всего несколько видов). Глубина обитания относительно небольшая, для большинства видов — 5-15 м, но некоторые виды распространены до глубины 40-100 м и даже 200 м. Бурые водоросли входят в экологическую группу бентосных (донных) организмов. Строение бурых водорослей Предшественниками хлоропластов бурых водорослей считаются бактерии, близкие к Heliobacterium chlorum. Основной фотосинтетический пигмент — хлорофилл а, вспомогательные — каратиноиды, в том числе бурый фукоксантин и желтые ксантофиллы. Вспомогательные пигменты бурых водорослей расширяют спектр поглощаемого ими света в сине-зеленой области. Запасное вещество - близкий к крахмалу растворимый углевод ламинарии. Слоевище (таллом) — только многоклеточное. Крупные, иногда многометровые слоевища бурых водорослей удерживаются на плаву благодаря расположенным в талломе воздушным пузырькам. У многих представителей бурых водорослей наметилась дифференцировка тканей. Внутри слоевища проходят сосудистые пучки, напоминающие флоэму высших растений. Появление сосудистой системы связано с необходимостью транспорта питательных веществ но многометровому слоевищу — от верхних, фотосинтезирующих, частей растения к нижним, у которых условия для фотосинтеза хуже. Размножение У бурых водорослей встречаются все формы полового размножения изогамия, гетерогамия и оогамия. Имеет место чередование поколений, обычно гетероморфных. Бесполое размножение — зооспорами и кусочками слоевища (вегетативное размножение). Значение бурых водорослей Бурые водоросли образуют на относительно небольшой глубине целые «подводные леса», окружающие сплошной стеной побережья всех морей и океанов обоих полушарий. Эти «подводные леса» служат источником пищи, укрытием и местом размножения для огромного числа морских обитателей, в том числе и для многих промысловых рыб. После отмирания водоросли образуют детрит, являющийся кормом для планктонных организмов. Бурые водоросли распространены повсеместно, однако самые крупные виды встречаются в морях умеренных и северных широт. К 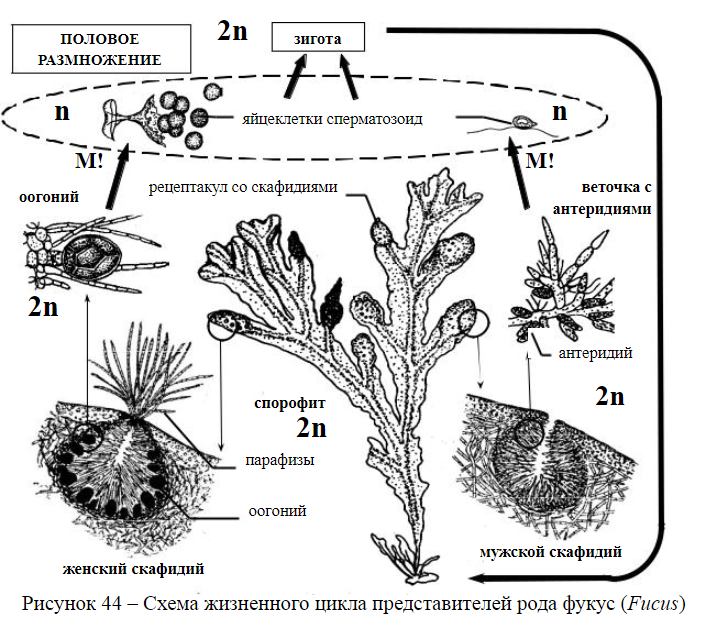 ласс: Бурые водоросли (Phaeophyceae) ласс: Бурые водоросли (Phaeophyceae)Порядок: Laminariales Семейство: Ламинариевые Род: Ламинария (Laminariaceae) Вид: Ламинария сахаристая (Laminaria saccharina) Вид:Фукус пузырчатый (Fucus vesiculosus). Несколько видов фукуса растут у нас в Балтийском и Северном морях. Они представляют собой дихотомически ветвящиеся бурые пластинки 0,5—1,0 м длиной и 1,5 см шириной. Водоросль диплоидна. На конечных разветвлениях таких пластинок в особых углублениях (скафидиях) развиваются половые органы — антеридии и оогонии, где при образовании сперматозоидов и яйцеклеток в их материнских клетках и происходит редукционное деление, и каждая материнская клетка дает гаметы. В антеридии образуется большое количество сперматозоидов, в оогонии — 8 яйцеклеток. Сперматозоиды по вскрытии антеридиев выплывают наружу (они подвижны, имея 2 жгутика); неподвижные яйцеклетки по вскрытии оогония вываливаются вводу, где (вне организма) и происходит слияние их в зиготу. Зигота прорастает в новую водоросль. У фукуса редукционное деление, отодвинуто на более позднюю стадию развития — к моменту образования половых элементов. Вся водоросль диплоидна. Смены поколений у фукуса нет. Есть смена ядерных фаз. 38.Типы организации таллома на примере представителей отдела Зелёные водоросли. Тело водорослей может быть одноклеточным, колониальным, многоклеточным. Структура таллома водорослей: Монадные водоросли (монады) представляют собой одноклеточные организмы со жгутиком. Они способны к передвижению в водной среде. Амебоидные (ризоподиальные) водоросли передвигаются за счет образования ложноножек (псевдоподий). Амебоидные клетки лишены клеточной стенки и способны постоянно менять свою форму. Одноклеточные водоросли, не имеющие ни жгутиков, ни псевдоподий и следовательно, неспособные к передвижению, называются коккоидными водорослями (шаровидной или округлой формы). Некоторые коккоидные водоросли прикрепляются к субстрату и выделяют большие количества слизи, окутывающую клетку толстым слоем. Это пальмеллоидная (капсанальная) структура таллома (пальмеллы). Как правило, пальмеллоидныеводоросли образуют колонии. Некоторые водоросли с другими структурами таллома (например, некоторые монадные и амебоидные) могут на время переходить в пальмеллоидное состояние. Простейшие многоклеточные водоросли имеют нитчатый (трихальный) таллом, в котором клетки соединены в цепочку друг за другом. Разнонитчатые (гетеротрихальные) водоросли отличаются от нитчатых тем, что образуют нити разных (двух или более) типов. Например, 11 некоторые водоросли имеют стелющиеся по субстрату и прямостоячие нити. Другие водоросли могут иметь крупную осевую и более мелкие боковые нити. Пластинчатые водоросли имеют таллом в виде пластинки, состоящей из одного или нескольких слоев клеток. Сифональные (сифоновые) водоросли – особая, редко встречающаяся структура таллома водорослей, при котором в пределах таллома отсутствуют клеточные перегородки. Такой таллом представляет собой как бы одну большую клетку с огромным количеством ядер. Сифонокладальная структура представлена многоядерными клетками, соединенными в нитчатые или иной формы многоклеточные талломы. Наиболее высокоорганизованная структура таллома – ложнотканевая (псевдопаренхиматозная). Такая структура напоминает пластинчатую, но состоит из клеток различных типов, образующих подобие тканей. У водорослей она встречается очень редко (только у бурых водорослей). Тканевая (паренхиматозная) структура образуется в результате делений клеток нити не только в поперечном, но и продольном направлении в виде паренхиматозных пластинок.  12 Рис. 3. Типы морфологической дифференциации таллома у водорослей: А — монадмый у Chlamydomonas; Б — амебоидный у Rhizochrysis; В — гсмимоиадмый у Hydrurus; Г — коккоидный у Pediastrum; Д — сарциноидный у Chlorosarcina; F — нитчатый у Ulothrix; Ж — разнонитчатый у Frilschiella; 3 — ложнотканевый у Furcellaria; И — тканевый у Laminaria; К — сифональный у Caulerpa; Л – сифонокладальный у Cladophora. 39. Отдел Папоротниковидные, особенности строения вегетативных и генеративных органов. Основные классы, семейства, представители. Представители этой группы широко распространены по земному шару, их можно встретить в разных климатических зонах и растительных сообществах. Предпочитают влажные местообитания, наиболее многочисленны в тропиках и субтропиках. Жизненные формы: преимущественно многолетние травянистые растения, реже деревья или лианы. Есть эпифитные (олений рог) и водные (сальвиния) формы. 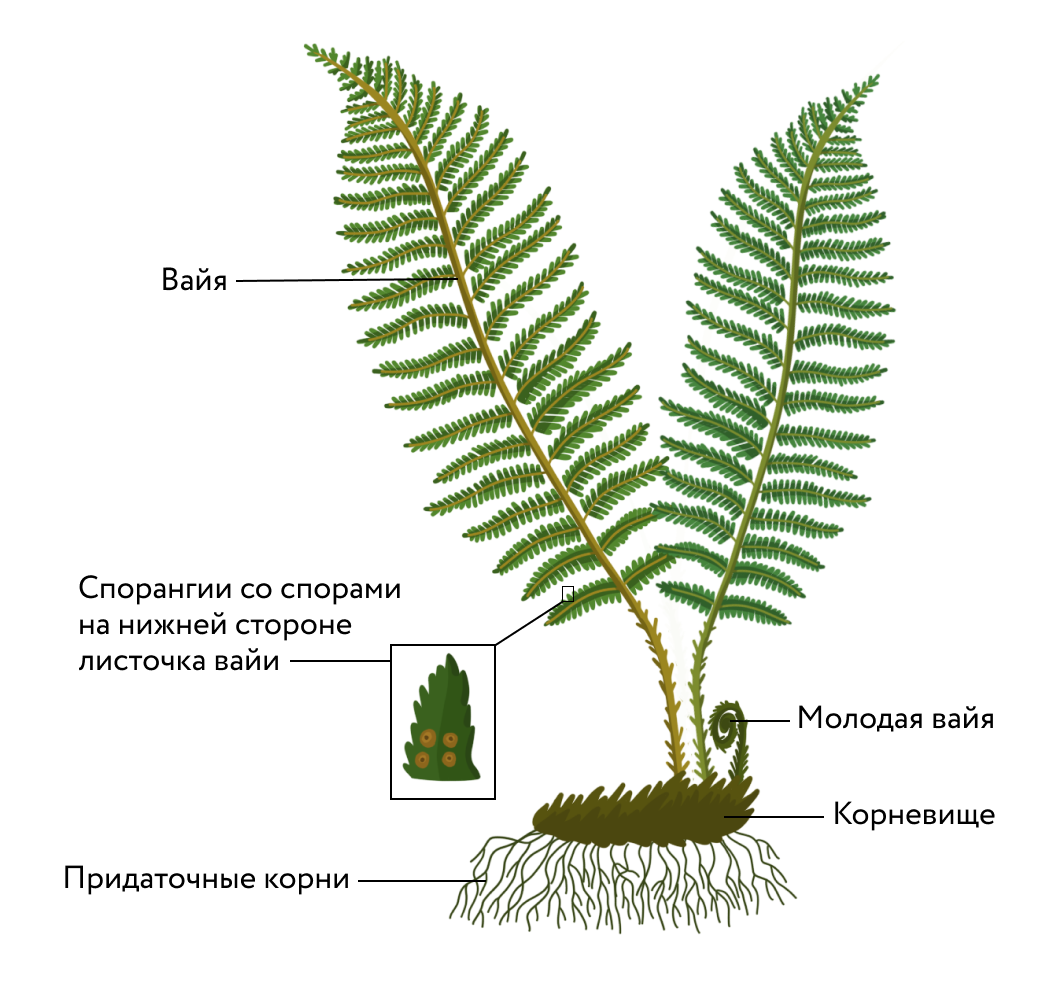 Самым крупным классом в отделе Папоротниковидные является класс Папоротниковые, или Настоящие папоротники. В лесах умеренной зоны нашей страны наиболее часто встречаются такие представители этого класса как щитовник мужской, кочедыжник женский, орляк, страусник. Самым крупным классом в отделе Папоротниковидные является класс Папоротниковые, или Настоящие папоротники. В лесах умеренной зоны нашей страны наиболее часто встречаются такие представители этого класса как щитовник мужской, кочедыжник женский, орляк, страусник.Строение Имеют вегетативные органы (придаточные корни и побег) и развитые ткани (покровные, механические, проводящие). У большинства папоротников умеренной зоны есть подземное корневище с придаточными корнями. На корневищах каждый год образуются новые розетки листьев. Рис. 2. Строение папоротника . Листья папоротников — вайи (от др.-греч. baion [ба́йон] — «пальмовая ветвь») — реже цельные, чаще сложно расчленённые, с хорошо развитой проводящей системой. В зачаточном состоянии вайя спирально свёрнута, её развитие происходит из этой «улитки». На обратной стороне или по краю листьев развиваются органы бесполого размножения — спорангии, в которых созревают споры.  Размножение РазмножениеПапоротники размножаются бесполым и половым путём. Бесполое размножение осуществляется спорами, а также вегетативно — укоренением побегов, листьев и, у некоторых видов, выводковыми почками, образующимися на листьях. Половое размножение происходит с участием половых клеток (гамет). Для передвижения сперматозоидов и успешного оплодотворения необходима капельно-жидкая влага. Особенности жизненного цикла В жизненном цикле папоротника чередуются бесполое (спорофит) и половое (гаметофит) поколения. На вайях образуются расположенные группами органы бесполого размножения — спорангии. Такие группы называются сорусами (от др.-греч. soros [сорос] — «куча»).  Рис. 4. Сорусы спорангиев на обратной стороне вайи папоротника Новая классификация папоротников построена на основе строения спорангиев и некоторых морфологических признаков. Это группа архегониальных споровых растений, состоит в отделе Polypodiophyta. Отдел представлен 10000 видами, которые объединены в 7 классов опсидов (сосудистые растения): К аневрофитам относят древнейшую примитивную группу. Археоптериды так же древнейшие папоротники, схожи с хвойными деревьями. Кладоксилы – слепая эволюционная ветвь. Класс Зигоптеридов представлен переходной группой. О фиоглоссы или Ужовники относятся к нынешним папоротникам. Маратти – папоротники-многолетники. К Полиподиям относятся многолетники и реже однолетники маленьких и больших размеров. Состоят из 3-х подклассов, чаще это равноспоровые растения.  В класс Полиподиопсидов входит подкласс Декоративных папоротников с комнатными, садовыми, скальными и аквариумными видами. Делят папоротники и по месту обитания. Наземные виды представлены травянистыми формами и стелющимися. На деревьях растут древесные формы (эпифиты), древовидные исполины (гигантская трава) вырастают вверх на 20-25м и в ширину до 50 см. Листья – по 2-3 м, а то и 5-6. Различить травяные и древовидные формы сложно. Редкие обитатели лесов – лиановидные папоротники, которые вырастают до 30 м. Воду и болотистую местность облюбовали водные папоротники. Комнатные виды: Род Адиантумов.Оригинальные курчавые папоротники, с перисто сложными, веерообразными нежными листьями на чёрно-коричневых черешках. Растения неприхотливые. Любители чаще интересуются видами: Венериным волосом, Стоповидным, Оленьим языком и Сколопендровым. Асплениумы. Эпифитные папоротники, у которых перисто рассечённые или цельные листья собраны розеткой. Распространённые виды асплениумов: у гнездовидного куст с цельными кожистыми листьями по 70 см; у живородящего вида – дговидно образные расчлененные вайи; у луковиценосного вида продолговато-треугольный лист. Полезный Нефролепсис.В роду насчитывается 40 видов выносливых наземных и эпифитных форм. Размножается не сложно горизонтальными побегами-усами. Очищает воздух в комнате и применяется для лечения как народное средство. Плавающие папоротники. Эти папоротники схожи с водорослями и относятся к водным растениям. Вместе с сальвинией сюда же относят марсилию, таиландского филипинса, азоллу и другие аквариумные виды. Страусовые перья. Этот вид, высотой 150 см, относится к роду Страусники, используется в садах чаще других папоротников. Высаживают растение в полутень, растёт на солнечных местах, возле прудов и на сырых почвах. Папоротник – водопад. Оригинальные резные вьющиеся лигодиумы. На одной стороне лианы растут цепкие корни, а на другой, в 2 ряда – перистые вайи светлого или тёмно-зелёного цвета 40. Принципы выделения отделов высших споровых растений. Сравнительная характеристика спорофита и гаметофита. Подцарство высших растений включает в себя зеленые растения, обитающие на суше. В зависимости от степени дифференцировки тканей и особенностей размножения их делят на — высшие споровые растения и семенные растения. Споровые растения размножаются и расселяются с помощью спор. Семенные растения морфологически более сложно устроены, и единицей размножения и расселения у них служит семя. В подцарстве высших известны миогочисленные ископаемые виды растений. Современные представители объединены в пять отделов споровых и два отдела семенных растений. Высшие споровые растения населяют сушу более 400 млн лет. Первые растения были небольшого размера, имели просто устроенные вегетативные органы. В процессе эволюции совершенствовалось внутреннее и внешнее строение. У высших споровых растений споры образуются в многоклеточных спорангиях и приспособлены к распространению ветром. Из спор развивается гаметофит, который называется заросток, на нем формируются половые органы. Для полового размножения необходима капельно-жидкая вода. Бесполое и половое поколения у споровых представлены самосто- ятельным и организмами. Отделы: моховидные, плауновидные, псилотовидные, хвощевидные, папоротниковидные. Отдел моховидные. К этой группе относятся древнейшие и наиболее просто устроенные высшие растения. Характерной чертой мхов, которая отличает их от всех высших растений, является доминирование в жизненном цикле гаметофита — полового поколения. Отдел плауновидные. К этой группе относятся растения с развитыми проводящими тканями. Вместе с хвощевидными и папоротниковидными они составляют группу сосудистых споровых растений. Отдел хвощевидные. Это самый малочисленный среди споровых растений отдел. В каменноугольном периоде хвощи были представлены большим разнообразием форм. Вымершие древовидные каламиты образовывали болотные леса. Современные хвощевидные — это многолетние травянистые растения. Отдел папоротниковидные.. В современном растительном покрове представители этого отдела самые распространенные из споровых растений. Более всего их во влажных тропиках. Там встречаются древовидные папоротники, высотой до 20 м, травянистые формы, эпифиты и лианы. Сравнительная характеристика гаметофита и спорофита. Сравнивая гаметофит и спорофит, приходится отметить, что они отличаются морфологически, физиологически к цитологически. Гаметофит, несомненно, преобладает в цикле развития мхов. Это объясняется тем, что гаметофит обладает автотрофным питанием и морфологически сложнее, тело его расчленено на органы, стебли и листья. Спорофит относительно проще по строению, не имеет вегетативных, органов и автотрофного питания, в своем существовании зависит от гаметофита. Вместе с тем в его организации лучше выражены признаки наземных растений — многослойность оболочки спорангия и спор, приспособления к рассеиванию спор. Чередование ядерных фаз у мхов соответствует чередованию поколений. Спорофит развивается из зиготы и является, следовательно, поколением диплоидным. Редукция происходит в археспории при образовании спор. Споры гаплоидны, гаплоидны также протонема и гаметофит. Таким образом, в цикле развития мхов явно преобладает гаплоидная фаза развития, что неблагоприятно для вида. Такое соотношение ядерных фах у других высших растений не встречается. Мхи не дали начала более прогрессивным линиям развития, они представляют боковую и тупую линию эволюции. 41. Образовательные ткани (меристемы), характеристика клеток их составляющих. Типы меристем. Второе название образовательной ткани растений – меристема. Слово происходит от греческого слова meristos – делимый. Основная особенность клеток меристем – постоянное деление, за счёт чего и происходит рост. Можно сказать, что эти клетки всё время или делятся, или готовятся к делению, накапливая энергию и нужные вещества. Строение клеток соответствует их деятельности. Клетки меристем мелкие, так как не успевают вырасти. Они имеют тонкие оболочки и крупные ядра. В цитоплазме много рибосом и митохондрий. Рибосомы синтезируют белковые молекулы для новых клеток. Митохондрии являются поставщиками энергии для разных клеточных процессов. Среди клеток меристемы есть два типа клеток: инициали; гистогены. Инициали выполняют только функцию деления и никогда не превращаются в клетки других тканей. Они способны делиться неопределённое число раз. Остальные клетки меристем называются гистогенами (от греческих слов histos – ткань, и genesis – происхождение). Они делятся несколько раз, а затем оттесняются новыми клетками и перестраиваются в клетки других тканей. Всё тело растения берёт начало от инициалей. Некоторые деревья благодаря наличию этих удивительных клеток продолжают свой рост на протяжении нескольких тысяч лет. Разделение на виды у меристем происходит по размещению в теле растения. Выделяют 4 вида меристем: верхушечные. Верхушечные меристемы расположены на верхушках корней и стеблей растения. При делении их клеток происходит рост корней вглубь, а стеблей вверх. боковые. Боковые меристемы (камбий) размещены в корне и стебле. На поперечном срезе имеют вид кольца. При делении их клеток идёт утолщение осевых органов (корня и стебля). вставочные. Вставочные или остаточные меристемы – это небольшие участки меристемы, оставшиеся от верхушечной в основании листьев. Они существуют временно и постепенно превращаются в другие ткани. раневые. Раневые меристемы образуются из других тканей в местах ранения растения. Они закрывают место ранения. 42. Анатомическое строение корня в зоне всасывания (на прмере корня ириса) и зоне проведения (на примере корня тыквы).  Рисунок 1 – Первичное строение корня IrisgermanicaL.: 1 – эпиблема; 2 – корневой волосок; 3 – экзодерма; 4 – мезодерма; 5 – эндодерма; 6 – пропускная клетка эндодермы; 7 – перицикл; 8 – луч ксилемы; 9 – участок флоэмы; 3–6 – первичная кора; 7–9 – центральный цилиндр Сравнительное анатомическое строение корня двудольных растений Корень вторичного строения двудольных растений имеет следующее строение: его поверхность покрывает пробка, состоящая из нескольких слоев омертвевших клеток, оболочки которых пропитаны суберином. Под нею находится феллоген – вторичная латеральная меристема, продуцирующая перидерму. Феллоген расположен над феллодермой, состоящей из нескольких слоев живых паренхимных клеток, формируемых феллогеном. Пробка, феллоген и феллодерма образуют комплекс вторичной покровной ткани – перидерму. Перидерма окружает проводящий цилиндр корня. Он состоит из нескольких проводящих пучков, разделенных между собой широкими лучами. Во внешней стороне пучков расположена флоэма, основной объем которой составляет вторичная флоэма. В ее наружном крае находятся узкие тангентальные прослойки первичной флоэмы, сформированные прокамбием. За вторичной флоэмой к оси корня следует пучковый камбий. Во внешнюю сторону камбий формирует новые слои клеток вторичной флоэмы и радиальных лучей. В сторону оси он продуцирует новые элементы вторичной ксилемы. Строение корня двудольных растений на примере корня тыквы обыкновенной (Cucurbita pepo L.) – постоянный препарат .  Рисунок 3 – Вторичное строение корня на примере корня Cucurbita pepo L.: 1 – вторичная флоэма; 2 – камбий; 3 – первичная ксилема; 4 – радиальный луч; 5 – сосуды вторичной ксилемы; 6 – ксилемная паренхима; 7 – основная паренхима вторичной коры; 8 – перидерма 43.Эволюционная оценка характерных признаков высших покрытосеменных растений и их различных жизненных форм. Наиболее крупный отдел растительного царства, насчитывающий более 350 семейств, 13 тыс. родов и до 240 тыс. видов. В настоящее время покрытосеменные распространены по всему земному шару — от тропиков до арктических и антарктических пустынь. Автотрофные цветковые растения — важнейший активный компонент биосферы. От их жизнедеятельности зависит течение общебиосферных процессов обмена веществ и трансформации энергии, газовый состав атмосферы, климат, водный-режим суши, характер процессов почвообразования. Понятно, что покрытосеменные определяют самую возможность существования большинства наземных животных. Они формируют среду обитания животного населения. Животные связаны с ними самыми разнообразными трофическими, консортивными и иными связями. Покрытосеменные растения отличаются необычайным полиморфизмом, удивительной эволюционной пластичностью и могут существовать в самых разнообразных условиях обитания, вплоть до безводных пустынь, скал, солончаков, разбитых песков, шахтных терриконов, трещин в асфальте или бетоне. Это единственная группа среди высших растений, представители которой сумели вторично освоить морскую среду: в соленых водах морских мелководий вместе с водорослями растут десятки видов покрытосеменных. Разнообразие размеров и жизненных форм цветковых растений, их приспособлений к конкретным условиям обитания поражает воображение. Самый крохотный их представитель, ряска волъфия (Wolffia arrhiza) едва достигает 1— 1,5 мм в диаметре и похожа на свободно плавающую зеленую водоросль, а попросту — на булавочную головку. С другой стороны, гигантские эвкалипты (виды рода Eucalyptus из семейства Myrtaceae) вырастают высотой до 100 м, а длина побегов тропических лиан — ротанговых пальм измеряется сотнями метром. Преобладают среди покрытосеменных автотрофные растения, но немало среди них полупаразитов, паразитов и сапротрофов, полностью утративших хлорофилл и способность к фотосинтезу. Эпифиты и эпифиллы (эпифиты, живущие на листьях других растений), деревья, кустарники, кустарнички, полукустарники и травы (многолетники и однолетники), плотоядные растения с их разнообразными приспособлениями для улавливания и переваривания мелких животных — это общеизвестные примеры. А тончайшие приспособления к опылению строго определенными агентами, к распространению плодов и семян, к перенесению неблагоприятных климатических периодов... Все это дает цветковым растениям возможность в полной мере реализовать свой эволюционно-адаптационный потенциал. Ни одна группа растений не могла выработать такого разнообразия приспособлений к определенным факторам среды. Признаки покрытосеменных Характернейшим признаком покрытосеменных (греч. antos — цветок, phyton — растение) считают цветок — отсюда и название Anthophyta. Покрытосеменные сначала приобрели совокупность действительно важнейших для них общих признаков — в первую очередь уникальные особенности жизненного цикла, а уже потом выработали разные цветки. При этом какого-то одного генерального направления эволюции не существовало, и в каждой эволюционной ветви покрытосеменных были осуществлены различные морфогенетические решения. Но при любых взглядах на цветок существенные его части — это андроцей, состоящий из одной, нескольких или многих тычинок (микроспорофиллов), и гинецей — из одного, нескольких или многих плодолистиков (мегаспорофиллов). Плодолистики могут быть свободными, и тогда каждый их них, срастаясь краями, образует пестик (один или несколько пестиков в цветке — по числу плодолистиков). При срастании нескольких плодолистиков формируется ценокарпный гинецей, пестик в нем всегда один. Важнейшая часть пестика — завязь. Именно наличие завязи как замкнутой полости, в которой развиваются семязачатки (в отличие от голосеменных, у которых семязачатки расположены на поверхности открытых мегаспорофиллов), обычно считают важнейшим признаком покрытосеменных, откуда и название — Angiospermae (греч. aggeion — сосуд, вместилище, sperma — семя). Близко по смыслу и название, предложенное в свое время И.Н. Горожанкиным, — Gynoeciatae (пестичные). Полагают, что завязь обеспечивает лучшую защиту семяпочек. Но это, конечно, не защита от животных-пожирателей: стенка завязи большей частью слишком тонка и нежна, чтобы устоять перед мощными жвалами жуков или грызущим аппаратом гусениц бабочек. Вряд ли завязь способна надежно предохранить семязачатки и от неблагоприятных внешних воздействий, в том числе от колебаний температуры, хотя в некоторых случаях эта цель, по-видимому, достигается. Вероятно, смысл прежде всего в том, что в полости завязи семязачатки оказываются в условиях влажной камеры, оптимальных для их развития на определенном этапе. Кроме того, благодаря образованию завязи расширяются и обогащаются возможности распространения зачатков: из цветка формируется плод, и теперь при диссеминации разлетаться и расселяться могут уже не только отдельные семена, но и целые плоды, у которых возникают разнообразные приспособления к распространению семян самым широким кругом агентов. Как и у некоторых голосеменных, у цветковых растений спермии доставлявются к яйцеклетке с помощью пыльцевой трубки — они «сифоногамны». Пыльцевая трубка обеспечивает высокую точность попадания спермиев в зародышевый мешок. Однако, коль скоро семязачатки скрыты в завязи и пыльца попасть непосредственно на микропиле не может, необходим какой-то специальный механизм, обеспечивающий улавливание пыльцы. Таким улавливающим аппаратом у покрытосеменных служит рыльце на верхушке столбика, или стилодия. Наличие рыльца — очень важная особенность. В свое время ван-Тигем даже предлагал называть покрытосеменные Stigmatae (рыльцевые, т.е. обладающие рыльцем). У всех архегониальных растений четко прослеживается эволюционная тенденция к разноспоровости, сопровождающаяся редукцией полового поколения в жизненном цикле. Покрытосеменные представляют собой как бы завершающий этап на этом пути. Предельное сокращение процессов микро- и мегаспорогенеза и ускорение развития гаметофитов, связанное с их крайней редукцией, были, вероятно, важными эволюционными факторами. В результате женский гаметофит цветковых растений представлен зародышевым мешком, состоящим в типе всего из 7 клеток, а мужской гаметофит — пылинкой, в которой всего 2 клетки — сифоногенная и спермагенная, дающая 2 спермия; при развитии мужского гаметофита нет даже следов про-таллиальных клеток. Одна из наиболее характерных особенностей покрытосеменных— двойное оплодотворение, при котором один из спермиев сливается с яйцеклеткой, образуя зиготу, а второй — с ядром центральной клетки зародышевого мешка, давая тем самым начало триплоидному эндоспер-м у. Это, по мнению ряда авторов, уникальный признак Angiospermae, аргумент в пользу их эволюционного единства и монофилетического происхождения. Часто высказываются соображения о возможности независимого и неоднократного возникновения процесса двойного оплодотворения в разных группах растений. Конечно, геометрически это можно себе представить, но с биологической и эволюционной точек зрения объяснение независимого в разных филах возникновения всей совокупности сложных механизмов, из которых складывается этот процесс, является дискуссионным. Одно из следствий двойного оплодотворения — формирование своеобразной питательной ткани, полиплоидного эндосперма (в отличие от гаплоидного эндосперма голосеменных, представляющего собой женский гаметофит). В деталях биологическое значение полиплоидного эндосперма не объяснено, но, по-видимому, он обеспечивает более благоприятные условия для развития зародыша и поэтому мог сыграть важную эволюционную роль. В вегетативной сфере подавляющее большинство покрытосеменных характеризуется наличием в ксилеме сосудов, тогда как почти у всех голосеменных древесина состоит только из трахеид. Лишь у немногих цветковых древесина гомоксильная, бессосудистая. Одни из них, как считают, сохранили этот анце-стральный признак, унаследовав его от предковых форм, а другие утратили сосуды в связи с существованием в особых экологических условиях, вроде водоемов, где подача воды по ксилеме большого значения не имеет. Вообще специализация тканей сыграла важную эволюционную роль. Паренхиматиза-ция позволила покрытосеменным выработать специальные запасающие ткани и разнообразные травянистые жизненные формы. В отличие от деревянистых растений у трав возможно сокращение онтогенеза и ускорение темпов эволюции; травянистые растения способны существовать в таких крайних условиях, в которых деревья и кустарники жить не могут. 44. Вторичные покровные ткани, их составляющие. Образование и строение чечевичек. Перидерма — вторичная комплексная покровная ткань. Она формируется на стеблях древесных растений к концу первого года жизни, покрывает многие подземные органы, изредка — плоды и другие части растений. Включает образовательную ткань феллоген, или пробковый камбий, и производные феллогена — пробку и феллодерму. Пробка, Или Феллема — многослойная, мертвая, плотная, опробковевшая (суберинизированная), водо - и газонепроницаемая защитная ткань. Феллодерма — живая, одно - или многослойная паренхимная ткань. Для водо - и газообмена в перидерме, под устьицами эпидермы из феллогена образуются Чечевички, Представляющие собой рыхлые участки, трещинки или вздутия. Чечевички функционируют в течение вегетационного периода, а на зиму закрываются слоем пробки, образованной феллогеном. Вторичное строение побега В  зоне первичной коры происходит заложение пробкового камбия, который начинает формировать пробку, в результате этого первичная кора оказывается снаружи от пробки и первичная покровная ткань эпидерма сбрасывается, замещается вторичной покровной тканью – перидермой. зоне первичной коры происходит заложение пробкового камбия, который начинает формировать пробку, в результате этого первичная кора оказывается снаружи от пробки и первичная покровная ткань эпидерма сбрасывается, замещается вторичной покровной тканью – перидермой.Клетки пробки пропитаны жироподобным веществом – суберином – и не пропускают воду и воздух, поэтому содержимое клетки отмирает, и она заполняется воздухом. Многослойная пробка образует своеобразный чехол стебля, надежно предохраняющий растение от неблагоприятных воздействий окружающей среды. Сосудистый камбий продолжает откладывать вторичные проводящие элементы, вплоть до наступления зимы. В таком виде, с вторичной покровной тканью перидермой, стебель зимует. Весной возобновляется деятельность сосудистого камбия, результатом чего являются видимые в ксилеме годичные кольца. Чечевички – это образования на стволе и ветках дерева, в которых находятся мелкие отверстия, прикрытые рыхлой тканью. По форме они очень разнообразны: обычно они представлены в виде мелких округлых бугорков, или штрихов, но могут быть и ромбическими. Основными функциями являются газообмен между внутренними живыми тканями стебля и окружающей средой, а также выведение лишней влаги. Чечевички разбросаны по стеблю и хорошо заметны, также их можно увидеть и на некоторых плодах в виде крапинок, например, на грушах, яблоках и др.  45. Механические ткани растений. Классификация. Значение механических тканей в жизни растений. Механические ткани в растении образуют своего рода «внутренний скелет», поддерживающий все остальные ткани и препятствующий их излому или разрыву. Выделяют два вида механических тканей: колленхима и склеренхима. Колленхима представлена в органах молодого растения и придает им упругость. У взрослых растений она сохраняется только в черешках листьев и обеспечивает их ориентацию в пространстве по отношению к солнцу. Колленхима состоит из живых клеток с неравномерно утолщенными оболочками, содержащими пектиновые вещества, которые способны набухать за счет поглощения воды. Со временем колленхима заменяется склеренхимой.  Склеренхима образована двумя видами мертвых клеток: волокнами (древесными и лубяными) и склереидами (каменистыми клетками). Волокна представлены длинными клетками с заостренными концами, стенки которых пропитаны лигнином. Древесные волокна входят в состав ксилемы у покрытосеменных и придают растениям прочность. У остальных растений они отсутствуют, и их функцию выполняют трахеиды. Лубяные волокна входят в состав флоэмы и придают растениям упругость. Стенки склереид пропитаны кремнеземом. Такие клетки встречаются в скорлупе орехов, в косточках (вишня, слива, абрикос) или в мякоти некоторых плодов. Механическая ткань образует каркас растительного организма, служит его опорой и придаёт прочность органам. Клетки этой ткани имеют сильно утолщённую клеточную стенку; они могут быть живыми или мёртвыми. Степень развития механических тканей различна у разных видов растений и зависит от условий обитания и размеров растения. Механические ткани слабо развиты у травянистых водных и прибрежных растений, но зато хорошо развиты у растений засушливых местообитаний и древесных форм. Клетки механической ткани присутствуют во всех органах растения, но наиболее многочисленны в стеблях, корнях, черешках и жилках листьев, оболочках семян и плодов. 46. Функции и классификация покровных тканей у растений. Строение эпидермы. Типы и строение устьиц. Все органы растения отделены от внешней среды покровными тканями. Покровные клетки плотно прилегают друг к другу и имеют прочные, иногда утолщённые оболочки. Межклеточного вещества почти нет. Основные функции покровных тканей — это защита растения от механических повреждений, высыхания, проникновения микроорганизмов, а также обеспечение газообмена и регулируемого испарения воды. Клетки покровных тканей могут быть живыми или мёртвыми. Листья, молодые зелёные стебли, цветки и многие плоды покрыты кожицей. Кожица (эпидерма) образована одним слоем живых клеток. В нём есть особые образования — устьица. В зависимости от условий окружающей среды устьица могут открываться, формируя отверстия, и закрываться. Через открытые устьица происходит испарение влаги и газообмен — перемещение кислорода и углекислого газа. Молодые корни тоже снаружи одеты покровной тканью, её называют эпиблемой или ризодермой. В эпиблеме корня нет устьиц, зато есть корневые волоски. Каждый корневой волосок — это длинная тонкостенная клетка покровной ткани. Соприкасаясь с частицами почвы, она всасывает воду с растворёнными в ней минеральными солями. На многолетних частях растения кожица замещается пробкой. Клетки пробки откладываются наружу и отмирают после созревания. Таким образом, слой пробки состоит преимущественно из мёртвых клеток. Эти клетки имеют толстую клеточную стенку, пропитанную жироподобным веществом, поэтому слой пробковых клеток практически не пропускает воду и воздух. Для обеспечения газообмена в пробке есть чечевички — бугорки, заполненные тонкостенными, рыхло расположенными округлыми клетками. Чечевички хорошо видны на молодых ветвях в виде бородавочек или штрихов. Новые слои клеток пробки откладываются регулярно. Поскольку внешний слой пробки состоит из мёртвых, не способных к растяжению клеток, он трескается. Наружная трещиноватая часть коры многолетних побегов и корней, состоящая из нескольких слоёв мёртвых клеток пробки и погибших элементов проводящей ткани, называется коркой. Корка, таким образом, состоит не только из клеток покровной ткани, поэтому её называют покровным комплексом. На старых стволах некоторых видов деревьев развивается очень мощная корка. Плантации пробкового дуба и амурского пробкового дерева (амурского бархата) выращивают ради того, чтобы ежегодно получать урожай корки. Из неё изготавливают пробки для винных бутылок, стеновые панели, покрытия для пола и многое другое. 47. Сущность и значение процессов мегаспорогенеза и образования зародышевого мешка. Мегаспоры образуются в процессе мегаспорогенеза. Процесс этот осуществляется в мегаспорангии, который у семенных растений называется нуцеллусом. Нуцеллус располагается в семязачатке, или семяпочке . У голосеменных на верхушке нуцеллуса закладываются архегонии , количество которых у разных групп различно. Каждый из архегониев содержит по одной крупной яйцеклетке. Однако, даже в случае успешного оплодотворения всех яйцеклеток, лишь одна из них развивается в семя. В мегаспорангии покрытосеменных только одна материнская клетка мегаспоры, имеющая, как и все растение, диплоидный набор хромосом, претерпевает дальнейшие изменения. Путем мейотического деления из нее возникают четыре гаплоидные, линейно расположенные мегаспоры. Обычно лишь одна из них продолжает развитие, а остальные отмирают. Эта клетка и формирует женский гаметофит. В типичном случае единственная мегаспора сильно увеличивается в размерах, а ее ядро делится несколько раз. Образовавшийся таким образом гаметофит , называемый зародышевым мешком , представляет собой обычно 8-ядерную клетку, у каждого из двух полюсов которой расположено по три ядра, а два ядра остаются в центре. Эти ядра, называемые полярными, чаще сливаются в диплоидное вторичное ядро. Далее в зародышевом мешке вокруг каждого из ядер формируются клеточные стенки. У верхушечного полюса зародышевого мешка образуются три клетки лицевого аппарата, одна из которых, обычно более крупная, функционирует как яйцеклетка , а две другие называются синергидами . Три клетки противоположного полюса получили название антипод. Синергиды и антиподы в процессе оплодотворения не участвуют и позднее дегенерируют. Образование зародышевого мешка В результате трёхкратного митотического деления ядра в полости зародышевого мешка образуются восемь ядер, которые облекаются цитоплазмой. Образуются лишённые оболочки клетки, которые располагаются в определённом порядке. На одном полюсе зародышевого мешка формируется яйцевой аппарат, состоящий из яйцеклетки и двух вспомогательных клеток. На противоположном полюсе располагаются три клетки (антиподы). С каждого полюса к центру зародышевого мешка мигрирует по одному ядру (полярные ядра). Иногда полярные ядра сливаются и образуют диплоидное центральное ядро зародышевого мешка. Зародышевый мешок, в котором произошла дифференцировка ядер, считается зрелым, он может воспринимать спермии.  К моменту созревания пыльцы и зародышевого мешка цветок раскрывается. 48.Цветок, его основные составляющие. Принципы составления формул и диаграмм цветков. Примеры цветков семейства Капустные (Brassicaceae), Бобовые (Fabaceae), Яснотковые (Lamiaceae), Лилейные (Liliaceae). Цветок состоит из цветоложа, чашечки, венчика, тычинок и пестика. |