ответы экзамен. Эволюция пищеварительной системы и особенности питания у представителей разных классов позвоночных животных
 Скачать 7.56 Mb. Скачать 7.56 Mb.
|

|
Условия дыхания животных на суше, органы и способы дыхания у дышащих воздухом животных. Успешная адаптация животных в процессе эволюции к воздушному дыханию осуществилась в широком масштабе только у членистоногих и позвоночных. Кроме того, хорошо приспособлены к жизни на суше некоторые улитки (они встречаются даже в пустынях) и сравнительно небольшое число других беспозвоночных. Дыхание животных в воздушной среде имеет ряд преимуществ по сравнению с таковым в воде. Во-первых, воздух отличается от воды гораздо более высоким содержанием кислорода и соответственно большей его доступностью для живых организмов. Так, 1 л воздуха содержит 210 мл О2, который весит 280 мг. Оставшаяся часть воздуха – 790 мл N2 – весит 910 мг. Таким образом, для того, чтобы экстрагировать из воздуха, пропускаемого через органы дыхания, нужное количество кислорода, необходимо пропустить в 3,5 раза большую часть воздуха, чем вес необходимого кислорода. 1 литр пресной воды, находящейся в равновесии с атмосферным воздухом, при температуре 15С содержит всего 7 мл кислорода. Эти 7 мл О2 весят 0,01 г (10 мг), что в 100 000 раз меньше веса воды. Таким образом, чтобы получить заданное количество кислорода, необходимо пропустить через дыхательные органы животного массу воды в 100 000 раз большую массы необходимого кислорода. Морская вода из-за присутствия солей характеризуется меньшей растворимостью газов, в связи с чем содержит еще меньше кислорода по сравнению с пресной (5,8 мл в 1 литре воды при температуре 15С). При повышении температуры воды растворимость газов в ней уменьшается, что обуславливает снижение содержания газов в ней. Так, при 30С пресная вода содержит 5,6 мл/л, а морская – 4,5 мл/л кислорода. Во-вторых, еще одно преимущество дыхания в воздухе – это сравнительно низкая его вязкость по сравнению с вязкостью воды. Так, вязкость воды при 20С равна 1 сПз (сантипаузу), а вязкость воздуха – 0,02 сПз (т.е. вязкость воды в 50 раз больше вязкости воздуха). Более низкая вязкость воздуха по сравнению с водой облегчает передвижение воздушных масс и делает возможным перемену направления движения воздуха в воздухоносных путях и органах дыхания в процессе дыхательного цикла. Вместе с тем, у некоторых позвоночных, дышащих воздухом (птиц и насекомых), встречается однонаправленное движение воздуха в органах дыхания. В водных дыхательных системах движение воды почти всегда однонаправленное, поскольку если бы поток воды входил и выходил в одном месте или двигался то туда, то сюда, то большую часть воды приходилось бы ускорять, затем останавливать, затем снова ускорять, но уже в обратном направлении. Для беспрерывных изменений кинетической энергии воды требовались бы большие затраты энергии, поэтому у всех водных животных, получающих кислород из воды, движение воды в органах дыхания осуществляется в одном направлении. В-третьих, скорость диффузии кислорода в воздушной среде примерно в 10 000 раз больше, чем в воде, при том же парциальном давлении (напряжении) кислорода. При такой быстрой диффузии размеры элементов дыхательного органа могут быть совсем иными, чем у животных, дышащих в воде. Так, расстояния, на которые газ диффундирует в легких, могут составлять несколько миллиметров, а диффузионные расстояния в жабрах рыб – ничтожные доли миллиметра. Несмотря на существующие преимущества дыхания животных в воздухе по сравнению с водной средой, существует и определенная трудность, связанная с испарением воды с поверхности воздухоносных путей и легких при дыхании. Вместе с тем, направленная внутрь сила для поглощения кислорода (159 мм рт. ст.) в 6 раз больше, чем сила для отдачи воды в сухой воздух (при 25С – 25 мм рт.ст.). Даже при высоких температурах (около 37С) давление водяных паров, обуславливающее отдачу воды в окружающую среду, мало по сравнению с давлением, заставляющим кислород поступать внутрь. Для того, чтобы не происходило чрезмерного испарения воды с дыхательной поверхности, наружный воздух не должен иметь к ней свободного доступа. В связи с этим газообменные поверхности обычно располагаются в специализированных дыхательных полостях (легких), что значительно ограничивает доступ воздуха. При этом обновление воздуха в такой полости часто очень тонко регулируется и не бывает более интенсивным, чем потребность их организма в кислороде. В связи с потерями воды вследствие испарения у наземных животных газообмен через всю поверхность тела встречается очень редко и, как правило, только во влажных местообитаниях. Например, дождевые черви, у которых весь дыхательный газообмен осуществляется через поверхность тела, очень чувствительны к недостатку воды. Они живут во влажных местах, и если остаются неприкрытыми на поверхности земли, то быстро высыхают и гибнут. Лучшая доступность кислорода в воздухе по сравнению с таковой в воде обусловила более высокий уровень метаболизма у них и в связи с этим более высокий уровень структурной и физиологической организации. Кроме того, гораздо более низкая теплопроводность воздуха по сравнению с теплопроводностью воды сделала возможным формирование в процессе эволюции гомойотермии (в связи с лучшими условиями по поддержанию на постоянном уровне температуры тела). В то же время некоторые очень крупные и быстро плавающие рыбы способны поддерживать температуру своего тела на более высоком уровне по сравнению с температурой воды. Органы дыхания у наземных животных У сухопутных животных встречаются 3 основных типа органов дыхания: легкие трахеи жабры. Жабры. Жабры довольно плохо приспособлены для дыхания в воздушной среде и используются лишь немногими животными – главным образом теми, которые сравнительно недавно перешли к сухопутному образу жизни и принесли с собой пережитки своего прежнего способа водного дыхания. Примером таких животных служат некоторые сухопутные крабы. Так, краб пальмовый вор (Birgus latro), почти полностью перешедший к наземной жизни (включая лазанье по кокосовым пальмам), имеет достаточно жесткие жабры, пригодные и для дыхания в воздухе. Другой сухопутный краб Cardiosoma может неопределенно долго жить и на суше и в воде, тогда как пальмовый вор через некоторое время погибает, если его держать погруженным в воду. Еще одна группа воздуходышащих ракообразных с жабрами – это наземные равноногие (Isopoda), называемые мокрицами. Эти животные предпочитают жить во влажных местах, и у видов, наиболее успешно приспособившихся к наземным местообитаниям, жабры заключены в полости, так что в функциональном плане их можно рассматривать как легкие. У некоторых рыб, способных дышать воздухом (но не у всех), функционирующие жабры сохранились. Так, обыкновенный угорь (Anguilla vulgaris) хорошо выживает на воздухе, если ему обеспечить достаточное охлаждение и влажность. Тогда большая часть кислорода поступает через кожу и меньшая – через жабры, поскольку жаберные нити частично слипаются, и в жаберной полости лишь небольшая часть их поверхности остается в контакте с воздухом. В результате угорь не получает обычного количества кислорода, и потребление кислорода уменьшается примерно вдвое. Легкие. Различают 2 типа легких: диффузионные и вентиляционные. Диффузионные легкие характеризуются тем, что обмен воздухом с окружающей средой осуществляется только путем диффузии. Легкие этого типа встречаются у относительно небольших животных, таких, как легочные улитки, скорпионы и некоторые равноногие раки. Диффузионные легкие у легочных улиток представляют собой часть мантийной полости, заполненную воздухом и имеющую густую капиллярную сеть. Такое легкое сообщается с окружающим воздухом через посредство единственного отверстия – пневмостома. Легкие скорпиона имеют вид мешков, представляющих собой углубления в теле паука, закрывающиеся особыми отверстиями-дыхальцами. Одна стенка у легочных мешков похожа на книжку: состоит из множества тончайших «складочек-страничек». В эти складочки затекает гемолимфа и через тоненькую стенку поглощает из воздуха легочных мешков кислород и отдает углекислый газ. Вентиляционные легкие имеются только у позвоночных. Значительное и регулярное обновление воздуха в легких необходимо при больших размерах тела, сочетающихся с высокой интенсивностью метаболизма. Дыхательные системы позвоночных вентилируются входящими и выходящими (приливно-отливными) потоками воздуха. У птиц эти системы намного сложнее, чем у млекопитающих, и устроены таким образом, что и при вдохе, и при выдохе воздух может течь через легкие в одном направлении. Трахеи. Этот тип органов дыхания характерен для насекомых. Он представляет собой систему трубочек, которые доставляют кислород непосредственно к тканям, так что циркуляция крови для транспорта газов оказывается ненужной. Газообмен в трахейной системе может происходить только путем диффузии, но у многих крупных и особенно высокоактивных насекомых имеет место и однонаправленное прокачивание воздуха через части трахейной системы. Преимущество однонаправленного потока состоит в том, что он обеспечивает намного лучший газообмен, чем при поочередном всасывании и выталкивании воздуха. Транспорт кислорода кровью. Влияние различных химических факторов и температуры на кривую диссоциации оксигемоглобина. Особенности кривых диссоциации оксигемоглобина у плодов млекопитающих в сравнении со взрослыми животными Возможны два основных способа транспорта кислорода кровью: в физически растворенном состоянии (непосредственно в транспортной жидкости, циркулирующей по сосудистой системе) в химически связанном со специальными дыхательными пигментами виде. При этом в состоянии физического растворения может переноситься очень малое количество кислорода (например, в крови млекопитающих концентрация кислорода в растворенном состоянии составляет около 0,2 мл на 100 мл крови) и поэтому такой способ доставки кислорода к тканям характерен лишь для очень мелких низкоорганизованных животных с низким уровнем метаболизма и соответственно низкой потребностью тканей в кислороде. У большинства же животных (в том числе, многих беспозвоночных, таких как ракообразные, паукообразные, моллюски и т.д.) для более эффективной доставки к тканям кислорода служат специальные дыхательные пигменты, находящиеся в свободном состоянии в плазме крови или в специальных клетках крови и способные обратимо химически связывать большие количества кислорода. Так, гемоглобин эритроцитов крови млекопитающих при максимальном насыщении обратимо связывает около 20 мл О2 на 100 мл крови, что в 100 раз больше количества физически растворенного в плазме крови кислорода. Таким образом, растворенный кислород имеет ничтожное значение по сравнению с кислородом, присоединенным к гемоглобину. Вещества, которые служат переносчиками кислорода в крови, — это белки, содержащие металл (обычно железо или медь). Они, как правило, окрашены, и поэтому их часто называют дыхательными пигментами. Самые распространенные из них следующие: гемоцианин – медьсодержащий белок, находящийся в свободном состоянии в плазме транспортирующей жидкости, гемоэритрин – железосодержащий белок непорфириновой структуры, всегда находится внутри клеток, хлорокруорин – железосодержащий белок, находящийся в растворе (плазме транспортирующей жидкости), гемоглобин – железосодержащий белок порфиринового типа, находится в растворе или внутри клеток; наиболее широко распространенный пигмент, типичен для позвоночных (почти всех, за исключением лептоцефалид (личинки угрей) и некоторых антарктических рыб (Chaenichthys)), и ряда беспозвоночных (иглокожих (голотурии), моллюсков, членистоногих, ракообразных,кольчатых червей, нематод, плоских червей (паразитические сосальщики), простейших, растений и грибов (дрожжи, Neurospora, корневые клубеньки бобовых растений (клевера, люцерны)). Дыхательные пигменты могут находиться в растворенном состоянии в плазме транспортирующей жидкости, или – заключены в специальные клетки (например, у позвоночных, тогда плазма крови не содержит растворенного пигмента). Упаковка дыхательных пигментов (в частности, гемоглобина) в специальные клетки (эритроциты), а не свободное нахождение его в растворенном состоянии в плазме крови дает несколько преимуществ. Первое возможное преимущество – уменьшение вязкости и коллоидно-осмотического давления плазмы. Второе важное преимущество, которое дает внутриклеточная локализация гемоглобина – это нахождение этого дыхательного пигмента в особой химической среде, создаваемой цитоплазмой эритроцита. Такая среда может по некоторым признакам существенно отличаться от плазмы крови и обеспечивать более эффективное связывание гемоглобином кислорода. На реакцию кислорода с гемоглобином, наряду с неорганическими ионами, сильно влияют и некоторые органические соединения, особенно органические фосфаты, и гемоглобин, локализованный внутри клеток, может быть обеспечен специфической, хорошо подобранной средой, отличной от плазмы. Кривые диссоциации оксигемоглобина у животных различных систематических групп и их зависимость от некоторых физико-химических факторов Наиболее важное свойство гемоглобина – это его способность обратимо связывать кислород: При больших концентрациях кислорода гемоглобин (НЬ) соединяется с кислородом, образуя оксигемоглобин (НbO2), и реакция идет в правую сторону. При низких концентрациях кислород отделяется, и реакция идет в левую сторону. Если концентрация О2 снижается до нуля, гемоглобин отдает весь связанный им кислород. Каждый атом железа в молекуле гемоглобина связывает одну молекулу кислорода, и когда при высокой концентрации (или высоком давлении) все имеющиеся места связывания заняты, гемоглобин становится полностью насыщенным и не может больше принимать кислород. При каждой данной концентрации кислорода существует определенное количественное соотношение (равновесие) между гемоглобином и оксигемоглобином. Если построить график зависимости количества оксигемоглобина от концентрации кислорода, получиться кривая кислородной диссоциации, которая описывает, каким образом эта реакция зависит от концентрации или парциального давления О2. На рис. приведена кривая диссоциации оксигемоглобина голубя. Она показывает, что кровь голубя полностью насыщается кислородом (100% НbО2) при парциальных давлениях О2 в альвеолярном воздухе 80-100 мм рт. ст. (10-13 кПа) и выше. При дальнейшем повышении давления кислорода гемоглобин больше не будет его связывать (т.е. достигается полное насыщение гемоглобина кислородом). При меньшем давлении О2 гемоглобин будет его отдавать, и при РО2 —30 мм рт. ст. (4 кПа) половина пигмента будет находиться в форме оксигемоглобина, а другая половина — в форме гемоглобина. Если давление О2 снижать дальше, все больше кислорода будет отделяться, пока при нулевом давлении О2 весь гемоглобин не будет полностью деоксигенирован. 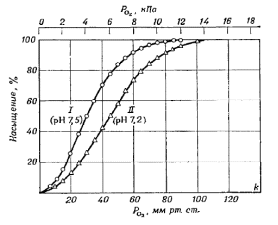 Рис. Кривые кислородной диссоциации для крови голубя. (Lutz at al., 1973.) I — кривая, полученная при нормальных для организма птицы условиях: температура 41°С, РСО2 35 мм рт. ст., рН 7,5; II — кривая, полученная после сдвига рН с 7,5 до 7,2 без изменения РCO2 У других животных кривые кислородной диссоциации гемоглобина по форме сходны с кривой голубя, но не идентичны ей. Кровь некоторых животных имеет более высокое сродство к кислороду (т.е. не так легко его отдает), и кривые диссоциации у них сдвинуты влево. Кровь других животных отдает кислород легче (ее сродство к кислороду меньше), и кривые диссоциации сдвинуты вправо. Наряду с различным сродством гемоглобина к кислороду, на связывание кислорода и тем самым на кривую диссоциации оксигемоглобина влияют и другие параметры (рН, ионная среда, органические фосфаты, температура). Эти различия имеют большое функциональное значение. Влияние температуры на связывание гемоглобином кислорода. Повышение температуры ослабляет связь между гемоглобином и кислородом, и эта связь легче разрывается. Поэтому при высоких температурах гемоглобин легче отдает кислород, и кривая диссоциации сдвигается вправо. С точки зрения физиологии это существенно, так как повышение температуры обычно вызвано увеличением интенсивности обмена веществ в ткани (а значит, и ее потребности в кислороде). Поэтому более легкая отдача О2 гемоглобином при повышенной температуре выгодна для организма. Это может иметь некоторое значение даже для теплокровных животных — например, в случаях лихорадки или при физической нагрузке, когда потребление кислорода возрастает. Влияние СО2 и рН на связывание гемоглобином кислорода. Существенное влияние на кривую диссоциации оказывает рН плазмы крови. Увеличение количества СО2 или других кислых продуктов метаболизма снижает рН плазмы, и это ведет к сдвигу кривой диссоциации вправо (рис.). Такой сдвиг означает, что повышенная концентрация СО2 приводит к повышенной отдаче кислорода при любом заданном его парциальном напряжении (давлении). Это явление известно как эффект Бора. В капиллярах тканей, где двуокись углерода поступает в кровь, гемоглобин отдает больше кислорода, чем это было бы при отсутствии влияния СО2 на его связывание. Следовательно, чем выше уровень метаболизма в тканях и соответственно чем больше СО2 и других кислых продуктов там образуется, тем больше кислорода отдает кровь (из-за понижения сродства гемоглобина к кислороду). Таким образом, можно сказать, что эффект Бора облегчает доставку тканям дополнительного количества кислорода. Эффект Бора не одинаково выражен у разных млекопитающих. Выраженность эффекта Бора зависит от размеров тела. В частности, гемоглобин мыши более чувствителен к закислению среды, чем гемоглобин слона. Отмеченная более высокая чувствительность гемоглобина к закислению у мелких животных важна для лучшей доставки кислорода к тканям мелких животных, у которых удельная интенсивность обмена выше, чем у более крупных животных и в связи с этим удельная потребность тканей в кислороде также выше. Влияние кислых продуктов метаболизма на кривую диссоциации гемоглобина у некоторых рыб выражено намного сильнее, чем у млекопитающих. Этот более сильный эффект называют эффектом Рута (рис.). Его самое важное следствие касается функции крови при секреции кислорода в плавательный пузырь. Для эффекта Рута характерно то, что при низком рН кровь не насыщается кислородом полностью ни при каком давлении О2. Это объясняется тем, что кровь у рыб содержит гемоглобин двух разных типов: высокочувствительный к кислоте и нечувствительный к кислоте. В присутствии высокой концентрации СО2 первый совсем не связывает кислород, тогда как второй дает обычную кривую диссоциации, характерную для этого вида гемоглобина. Зачем рыбам два различных гемоглобина? Легко понять значение молекул Нb, чувствительных к кислоте, но для чего нужна другая разновидность, которая к кислоте нечувствительна? Какая в этом необходимость? Дело, очевидно, в том, что у рыбы, когда она в критической ситуации развивает при плавании максимальные усилия, образуются большие количества молочной кислоты. Если бы в присутствии этой кислоты весь гемоглобин терял способность связывать кислород, рыба погибла бы от асфиксии. Однако наличие другого, нечувствительного к кислоте гемоглобина предотвращает эту беду. Влияние органических фосфатов на связывание гемоглобином кислорода. Перенос газов эритроцитами тесно связан с метаболизмом самих эритроцитов. Эритроциты млекопитающих не имеют ядра, и в течение многих лет их рассматривали как нечто мало отличающееся от инертного мешка, набитого гемоглобином. Однако в эритроците имеет место активный углеводный обмен, который не только существен для жизнеспособности клетки, но и оказывает глубокое влияние на ее функцию при транспорте кислорода. Эритроцитам свойственно также высокое содержание аденозинтрифосфата (АТФ) и еще более высокий уровень 2,3-дифосфоглицерата (ДФГ). Присутствие этих органических фосфатов существенно влияет на сродство гемоглобина к кислороду и помогает объяснить, почему раствор чистого гемоглобина проявляет гораздо большее сродство к кислороду, чем цельная кровь (т.е., когда гемоглобин находится в специальной среде, создаваемой цитоплазмой эритроцитов). В частности, кривая диссоциации для Нb в растворе сдвинута далеко влево по отношению к кривой диссоциации оксигемоглобина цельной крови (раствор гемоглобина характеризуется гораздо более высоким сродством к кислороду, чем гемоглобин эритроцитов). Если к раствору гемоглобина добавить органические фосфаты, в особенности ДФГ, его сродство к кислороду сильно снижается и становится близким к уровню интактных клеток. Это снижение можно объяснить тем, что к гемоглобину присоединяется ДФГ, который изменяет его сродство к О2. Открытие влияния ДФГ на способность гемоглобина присоединять кислород объясняет многие наблюдения, которые раньше казались совсем непонятными. Оно также объясняет, почему при исследовании цельной крови можно наблюдать ряд особенностей кривой диссоциации, которые не выявляются при изучении гемоглобина в растворе. Так, например, у высокогорных жителей уровень ДФГ в эритроцитах выше, чем у людей, живущих на уровне моря. Вследствие этого на больших высотах кривая диссоциации О2 сдвинута вправо (сродство гемоглобина к кислороду понижено, что обуславливает лучшую отдачу кислорода тканям). Этот сдвиг был хорошо известен, но механизм его оставался неясным до тех пор, пока не было установлено, что уровень ДФГ возрастает в ответ на гипоксию. Влияние ДФГ и других фосфатов на сродство гемоглобина к кислороду позволило объяснить давно известный факт, что при хранении консервированной крови сродство ее к кислороду со временем увеличивается. Оно также помогает понять, почему очищенные, освобожденные от ДФГ гемоглобины плода и взрослого человека обнаруживают одинаковое сродство к кислороду, хотя, у крови плода сродство к кислороду выше. Кроме того, теперь можно объяснить сдвиг вправо кривых диссоциации в период беременности, хотя беременная самка (в отличие от плода) все время имеет один и тот же гемоглобин (НbА, проявляющий более низкое сродство к кислороду, чем фетальный гемоглобин плода). Дело в том, что в период беременности в эритроцитах матери возрастает содержание ДФГ, что обуславливает понижение сродства гемоглобина к кислороду и соответственно лучшую его отдачу крови плода, в эритроцитах которого присутствует фетальный гемоглобин, проявляющий более высокое сродство к кислороду чем антильный гемоглобин матери. |