ответы экзамен. Эволюция пищеварительной системы и особенности питания у представителей разных классов позвоночных животных
 Скачать 7.56 Mb. Скачать 7.56 Mb.
|

|
Продуценты — это живые организмы, которые способны синтезировать органическое вещество из неорганических составляющих с использованием внешних источников энергии. (Отметим, что получение энергии извне — общее условие жизнедеятельности всех организмов; по энергии все биологические системы — открытые.) Их называют также автотрофами, поскольку они сами снабжают себя органическим веществом. В природных сообществах продуценты выполняют функцию производителей органического вещества, накапливаемого в тканях этих организмов. Органическое вещество служит и источником энергии для процессов жизнедеятельности; внешняя энергия используется лишь для первичного синтеза. Все продуценты по характеру источника энергии для синтеза органических веществ подразделяются на фотоавтотрофов и хемоавто- трофов. Первые используют для синтеза энергию солнечного излучения в части спектра с длиной волны 380—710 нм. Это главным образом зеленые (хлорофиллоносные) растения, но к фотосинтезу способны и представители некоторых других царств органического мира. Среди них особое значение имеют цианобактерии (синезеленые «водоросли»), которые, по-видимому, были первыми фотосинтетиками в эволюции жизни на Земле. Способны к фотосинтезу также многие бактерии, которые, правда, используют особый пигмент — бактериох- лорин — и не выделяют при фотосинтезе кислород. Основные исходные вещества, используемые для фотосинтеза, — диоксид углерода и вода (основа для синтеза углеводов), а также азот, фосфор, калий и другие элементы минерального питания. Создавая органические вещества на основе фотосинтеза, фотоавто- трофы, таким образом, связывают использованную солнечную энергию, как бы запасая ее. Последующее разрушение химических связей ведет к высвобождению такой «запасенной» энергии. Это относится не только к использованию органического топлива; «запасенная» в тканях растений энергия передается в виде пищи по трофическим цепям и служит основой потоков энергии, сопровождающих биогенный круговорот веществ. Хемоавтотрофы в процессах синтеза органического вещества используют энергию химических связей. К этой группе относятся только прокариоты: бактерии, архебактерии и отчасти синезеленые. Химическая энергия высвобождается в процессах окисления минеральных веществ. Экзотермические окислительные процессы используются нитрифицирующими бактериями (окисляют аммиак до нитритов, а затем до нитратов), железобактериями (окисление закисного железа до окис- ного), серобактериями (сероводород до сульфатов). Как субстрат для окисления используется также метан, СО и некоторые другие вещества. При всем многообразии конкретных форм продуцентов-автотро- фов их общая биосферная функция едина и заключается в вовлечении элементов неживой природы в состав тканей организмов и таким образом в общий биологический круговорот. Суммарная масса автотро- фов-продуцентов составляет более 95 % массы всех живых организмов в биосфере. Консументы - живые существа, не способные строить свое тело на базе использования неорганических веществ, требующие поступления органического вещества извне, в составе пищи, относятся к группе гетеротрофных: организмов, живущих за счет продуктов, синтезированных фото- или хемосинтетиками. Пища, извлекаемая тем или иным способом из внешней среды, используется гетеротрофами на построение собственного тела и как источник энергии для различных форм жизнедеятельности. Таким образом, гетеротрофы используют энергию, запасенную автотрофами в виде химических связей синтезированных ими органических веществ. В потоке веществ по ходу круговорота они занимают уровень потребителей[1], облигатно связанных с автотрофными организмами (консументы I порядка) или с другими гетеротро- фами, которыми они питаются (консументы II порядка; рис. 2.1). 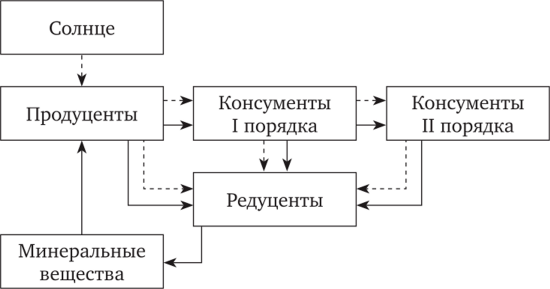 Рис. 2.1. Упрощенная схема переноса вещества (сплошная линия) и энергии (пунктирная линия) в процессе биологического круговорота (по В. Е. Соколову, И. А. Шилову, 1989) К консументам относится огромное количество живых организмов из разных таксонов. Их нет лишь среди цианобактерий и водорослей. Из высших растений к консументам относятся бесхлорофилльные формы, паразитирующие на других растениях; частично положение консументов занимают и растения со смешанным питанием (например, насекомоядные типа росянки). Все животные — консументы, и их роль в поддержании устойчивого биогенного круговорота очень велика. Общее значение консументов в круговороте веществ своеобразно и неоднозначно. Они не обязательны в прямом процессе круговорота: искусственные замкнутые модельные системы, составленные из зеленых растений и почвенных микроорганизмов, при наличии влаги и минеральных солей могут существовать неопределенно долгое время за счет фотосинтеза, деструкции растительных остатков и вовлечения высвобожденных элементов в новый круговорот. Но это возможно лишь в стабильных лабораторных условиях. В природной обстановке возрастает вероятность гибели таких простых систем от многих причин. «Гарантами» устойчивости круговорота и оказываются в первую очередь консументы. В процессе собственного метаболизма гетеротрофы разлагают полученные в составе пищи органические вещества и на этой основе строят вещества собственного тела. Трансформация первично продуцированных автотрофами веществ в организмах консументов ведет к увеличению разнообразия живого вещества. Разнообразие же — необходимое условие устойчивости любой кибернетической системы на фоне внешних и внутренних возмущений (принцип Эшби). Живые системы — от организма до биосферы в целом — функционируют по кибернетическому принципу обратных связей. В дальнейшем тексте мы не раз встретимся с важностью различных форм биологического разнообразия (биологической разнокачественности) для устойчивого функционирования экосистем[2]. Животные, составляющие основную часть организмов-консументов, отличаются подвижностью, способностью к активному перемещению в пространстве. Этим они эффективно участвуют в миграции живого вещества, дисперсии его по поверхности планеты, что, с одной стороны, стимулирует пространственное расселение жизни, а с другой — служит своеобразным «гарантийным механизмом» на случай уничтожения жизни в каком-либо месте в силу тех или иных причин. Примером такой «пространственной гарантии» может служить широко известная катастрофа на о. Кракатау: в результате извержения вулкана в 1883 г. жизнь на острове была полностью уничтожена, но в течение всего 50 лет восстановилась — было зарегистрировано порядка 1200 видов. Заселение шло главным образом за счет не затронутых извержением Явы, Суматры и соседних островов, откуда разными путями растения и животные вновь заселили покрытый пеплом и застывшими потоками лавы остров. При этом первыми (уже через 3 года) на вулканическом туфе и пепле появились пленки цианобактерий. Процесс становления устойчивых сообществ на острове продолжается; лесные ценозы еще находятся на ранних стадиях сукцессии и сильно упрощены по структуре. Наконец, чрезвычайно важна роль консументов, в первую очередь животных, как регуляторов интенсивности потоков вещества и энергии по трофическим цепям. Способность к активной авторегуляции биомассы и темпов ее изменения на уровне экосистем и популяций отдельных видов в конечном итоге реализуется в виде поддержания соответствия темпов создания и разрушения органического вещества в глобальных системах круговорота. Участвуют в такой регуляторной системе не только консументы, но последние (особенно животные) отличаются наиболее активной и быстрой реакцией на любые возмущения баланса биомассы смежных трофических уровней. Подробнее регуляторные механизмы в популяциях и экосистемах будут рассмотрены ниже. В принципе система регулирования потоков вещества в биогенном круговороте, основанная на комплементарности составляющих эту систему экологических категорий живых организмов, работает по принципу безотходного производства. Однако в идеале этот принцип соблюден быть не может в силу большой сложности взаимодействующих процессов и влияющих на них факторов. Результатом нарушения полноты круговорота явились отложения нефти, каменного угля, торфа, сапропелей. Все эта вещества несут, в себе энергию, первоначально запасенную в процессе фотосинтеза. Использование их человеком — как бы «отставленное во времени» завершение циклов биологического круговорота. Редуценты - к этой экологической категории относятся организмы- гетеротрофы, которые, используя в качестве пищи мертвое органическое вещество (трупы, фекалии, растительный опад и пр.), в процессе метаболизма разлагают его до неорганических составляющих. Частично минерализация органических веществ идет у всех живых организмов. Так, в процессе дыхания выделяется С02, из организма выводятся вода, минеральные соли, аммиак и т. д. Истинными редуцентами, завершающими цикл разрушения органических веществ, следует поэтому считать лишь такие организмы, которые выделяют во внешнюю среду только неорганические вещества, готовые к вовлечению в новый цикл. В категорию редуцентов входят многие виды бактерий и грибов. По характеру метаболизма это организмы-восстановители. Так, денитрифицирующие бактерии восстанавливают азот до элементарного состояния, сульфатредуцирующие бактерии — серу до сероводорода. Конечные продукты разложения органических веществ — диоксид углерода, вода, аммиак, минеральные соли. В анаэробных условиях разложение идет дальше — до водорода; образуются также углеводороды. Полный цикл редукции органического вещества более сложен и вовлекает большее число участников. Он состоит из ряда последовательных звеньев, в череде которых разные организмы-разрушители поэтапно превращают органические вещества сначала в более простое формы и только после этого в неорганические составляющие действием бактерий и грибов. Учение о биосфере Неклеточные формы жизни в биосфере: вирусы. Роль вирусов в биосфере Земли. Ви́рус (от лат. vira) — микроскопическая частица, состоящая из белков и нуклеиновых кислот и способная инфицировать клетки живых организмов. Вирусы являются облигатными паразитами — они не способны размножаться вне клетки. В настоящее время известны вирусы, размножающиеся в клетках растений, животных, грибов и бактерий (последних обычно называют бактериофагами). Обнаружен также вирус, поражающий другие вирусы. Вирусы представляют собой молекулы нуклеиновых кислот (ДНК или РНК), заключённые в защитную белковую оболочку (капсид). Наличие капсида отличает вирусы от других инфекционных агентов, вироидов. Вирусы содержат только один тип нуклеиновой кислоты: либо ДНК, либо РНК. Ранее к вирусам также ошибочно относили прионы, однако впоследствии оказалось, что эти возбудители представляют собой особые белки и не содержат нуклеиновых кислот. Вирусы являются одной из самых распространённых форм существования органической материи на планете по численности: воды мирового океана содержат колоссальное количество бактериофагов (около 250 миллионов частиц на миллилитр воды), их общая численность в океане — около 4·1030а численность вирусов (бактериофагов) в донных отложениях океана практически не зависит от глубины и всюду очень высока. В океане обитают сотни тысяч видов (штаммов) вирусов, подавляющее большинство которых не описаны и тем более не изучены. Вирусы играют важную роль в регуляции численности популяций животных. Сравнительная физиология животных Характер тока воды и крови в жабрах рыб и его физиологическое значение. Механизмы, обеспечивающие поддержание тока воды в жабрах рыб. У рыб жабры имеют сложное строение, обеспечивающее увеличение площади их респираторной поверхности, а также противоток воды и крови в жаберных пластинках. Кроме того, жабры рыб заключены в жаберную полость, обеспечивающую защиту крупных органов и позволяющую обтекать жаберную поверхность наиболее эффективным образом. Строение жабр рыб. Жаберные щели рыб разделены жаберными дугами). От каждой жаберной дуги отходят два ряда жаберных нитей (или жаберных перегородок). Кончики нитей, соприкасаются, заставляя воду течь между нитями. На каждой жаберной нити располагаются ряды плотно уложенных жаберных пластинок (или лепестков). Газообмен между водой и кровью осуществляется в жаберных пластинках (дискообразной формы). Жаберные пластинки свисают в жаберную полость. Жаберная полость снаружи прикрыта кожной складкой, укреплённой костями,— жаберной крышкой. Т.о респираторная поверхность жабр рыб значительно увеличена за счёт тончайших складок на поверхности жаберных пластинок. Поток воды около жабры и поток крови внутри этого органа направлены противоположно друг другу. Принцип противотока имеет важное значение для эффективной экстракции кислорода из омывающей жабру воды. Так, когда кровь уже готова покинуть жаберную пластинку, она встречает воду, только что вошедшую между жаберными пластинками. Таким образом, эта кровь поглощает кислород из воды, в которой содержание кислорода еще не понизилось и в результате насыщение крови кислородом может достигнуть максимального уровня. По мере дальнейшего прохождения между пластинками вода встречает кровь со все более и более низким содержанием кислорода, и поэтому она будет все время отдавать кислород. Благодаря отмеченному противотоку в жабрах жаберные пластинки извлекают кислород из воды по всей длине, и вода, пройдя жабру, может потерять до 80-90% исходного количества кислорода. Это очень высокая степень экстракции кислорода (для сравнения в легких млекопитающих из альвеолярного воздуха экстрагируется около 25% кислорода). Механизмы, обеспечивающие поддержание тока воды в оперкулярных полостях. Для того чтобы жабры эффективно извлекали из омывающей их воды кислород, необходимы: постоянная смена воды у дыхательных поверхностей жабр (поскольку в отсутствии движения воды парциальное напряжение кислорода в ней у дыхательной поверхности жабры быстро снизиться вплоть до нуля и дальнейший газообмен будет фактически не эффективен) тесный контакт между водой и жаброй. Для создания тока воды над поверхностью жабр существуют различные механические приспособления, способные обеспечивать движение воды 2-мя путями: движением жабры в воде перемещением воды вокруг жабры. Движение жабры в воде «практично» только для мелких организмов. У рыб реализуется 2-й механизм – движение воды над дыхательной поверхностью жабры. Такое движение у рыб создается благодаря механическим приспособлениям, подобным насосу. Эта система для прокачивания воды через жабры состоит из 2-х насосов: ротовой полости и оперкулярной полости. Увеличение объема первого насоса (ротовой полости) достигается путем опускания челюсти, а увеличение объема оперкулярной полости (расположены по одной с каждой стороны, сообщаются с ротовой полостью) – за счет движения жаберных крышек при одновременном блокировании обратного тока воды вблизи краев кожными щитками, играющими роль пассивных клапанов. Действие этих 2-х насосов таково, что ток воды через жабры поддерживается на протяжении почти всего дыхательного цикла. Вода продолжает течь, несмотря на то, что во время части цикла давление в ротовой полости может сделаться меньше, чем во внешней среде, но при этом давление в оперкулярной полости поддерживается на еще более низком уровне, чем во рту. Таким образом, давление в ротовой полости на протяжении почти всего дыхательного цикла выше такового в оперкулярных полостях, что обеспечивает практически непрерывный ток воды через жабры. Некоторые рыбы не способны дышать таким способом. Для этих рыб характерен другой механизм поддержания постоянного тока воды вокруг жабр – таранная вентиляция. Сущность ее состоит в том, что они плавают с постоянно приоткрытым ртом. При этом видимые дыхательные движения у них отсутствуют, но в связи с непрерывным плаванием с открытым ртом вода все равно непрерывно течет через жабры. Таранная вентиляция встречается не только у крупных, быстро плавающих пелагических рыб. Многие рыбы при малых скоростях используют для дыхания прокачивание, а при увеличении скорости переходят к таранной вентиляции. При этом переход к таранной вентиляции наблюдается в диапазоне скоростей от 0,5 до 1 м/с: при больших скоростях активные дыхательные движения прекращаются. Переход к таранной вентиляции не означает, что дыхание «пускается на самотек»; это значит только, что вентиляцию обеспечивают теперь не мышцы оперкулярного насоса, а плавательные мышцы тела и хвоста. При этом открытый рот увеличивает лобовое сопротивление, которое должно быть компенсировано усиленной работой мускулатуры. Однако возможно, что при высокой интенсивности дыхания, необходимой во время быстрого плавания, непрерывный ток воды при таранной вентиляции энергетически более выгоден, чем оперкулярное прокачивание. Степень открытия рта регулируется таким образом, чтобы обеспечивать достаточный ток воды через жабры, но не увеличивать лобовое сопротивление больше, чем это требуется для адекватной вентиляции. Если при протекании воды через жабры в них застревают твердые взвешенные частицы, то рыбы удаляют их за счет изменения направления тока воды. В частности, они расширяют ротовую полость (путем опускания нижней челюсти) при сомкнутых губах, что приводит к резкому снижению давления во рту и, как следствие, движению воды из оперкулярных полостей в ротовую полость (т.е. в обратном обычному направлении). Данный механизм аналогичен кашлю у млекопитающих, служащему для удаления слизи и твердых частиц из нижних дыхательных путей. Общий план строения и функционирования дыхательной системы птиц. Сущность и физиологическая роль двойного дыхания птиц. Органы дыхания у птиц сильно отличаются от соответствующих органов млекопитающих. В частности, у птиц имеются небольшие компактные легкие, которые сообщаются с объемистыми тонкостенными воздушными мешками – воздушными пространствами, расположенными между внутренними органами и даже разветвляющимися внутри костей черепа и конечностей. Эта сложная дыхательная система отчасти облегчает тело птицы (а именно, такую функцию выполняют воздушные мешки, заполняющие полости костей) и дает некоторые преимущества в плане увеличения насыщения крови кислородом при полете, когда из-за интенсификации метаболизма потребность в кислороде существенно возрастает (в 8-10 раз по сравнению с его потреблением в покое). Вместе с тем, дыхательная система птиц не является определяющей для способности их к полету, поскольку такой способностью обладают и некоторые млекопитающие (летучие мыши), не имеющие воздушных мешков. Кроме того, 8-10-ти кратное увеличение потребления организмом кислорода при интенсификации энергозатрат возможно и у млекопитающих. Объем легких типичной птицы составляет чуть больше половины объема легких млекопитающего той же величины (например, у птицы весом 1 кг – 29,6 мл, тогда как у млекопитающего такой же массы – 53,5 мл). Напротив объем трахеи у птиц (3,7 мл) намного больше, чем у млекопитающих (0,9 мл), что отчасти связано с большей длиной шеи у птиц. Вместе с тем, малый объем легких птиц с лихвой компенсируется воздушными мешками, по объему в несколько раз превышающими легкие (у птицы весом 1 кг воздушные мешки имеют объем 127,5 мл), тогда как у млекопитающих воздушных мешков нет совсем. Благодаря наличию воздушных мешков общий объем дыхательной системы у птиц примерно в 3 раза больше, чем у млекопитающих соответствующих размеров (например, у птицы весом 1 кг общий объем дыхательной системы составляет 160,8 мл, тогда как у млекопитающего аналогичной массы – 54,4 мл). Различие между птицами и млекопитающими не ограничивается только воздушными мешками; сами легкие птиц по своему строению тоже радикально отличаются от легких млекопитающих. В частности, у млекопитающих самые тонкие веточки бронхов заканчиваются мешковидными альвеолами, тогда как у птиц тончайшие разветвления бронхиальной системы (называемые парабронхами) допускают сквозной проход воздуха. Таким образом, у птиц воздух может течь через легкие непрерывным потоком мимо газообменной поверхности воздушных капилляров (тонкостенные густо оплетенные капиллярами выросты слизистой парабронхов); у млекопитающих же воздух должен входить в альвеолы и выходить из них. Это наиболее важное различие между дыхательными системами птиц и млекопитающих, и из него вытекают глубокие физиологические следствия. Анатомически и функционально воздушные мешки образуют две группы: заднюю, включающую обширные брюшные мешки, и переднюю, которая состоит из нескольких мешков меньшего размера. Аналогично млекопитающим, трахея птиц делится на два бронха, каждый из которых входит в соответствующее легкое, но затем, в отличие от млекопитающих, главный бронх в буквальном смысле проходит сквозь легкое и заканчивается в брюшном мешке. Передние мешки присоединяются к этому главному бронху в передней части легких; задние мешки присоединены к заднему отделу главного бронха. Главный бронх сообщается также с легкими, и, кроме того, некоторые из воздушных мешков непосредственно связаны с легочной тканью. Стенки воздушных мешков тонки, непрочны, слабо васкуляризированы и не имеют ни складок, ни гребней, которые увеличивали бы поверхность. В отличие от альвеол легких они не принимают участия в газообмене с кровью, а, вероятнее всего, подобно мехам, служат для втягивания и выталкивания воздуха. В частности, во время вдоха по мере того, как воздушные мешки расширяются, в них понижается давление (одновременно и в передних, и в задних мешках), и воздух входит во все мешки (но при этом не все мешки наполняются именно наружным воздухом). Наружный воздух через главный бронх при вдохе поступает в задние мешки, тогда как передние наполняются воздухом, выталкиваемым из легких. Таким образом, передние мешки служат резервуарами для приема из легких того воздуха, который должен быть выведен наружу при следующем выдохе. Во время выдоха давление в мешках увеличивается, и воздух из них снова выходит. Но при этом, воздух из задних мешков не выходит через главный бронх наружу, а направляется в легкие. При следующем вдохе воздух из легких переходит в передние мешки, и, наконец, при втором выдохе воздух из передних мешков выходит прямо наружу. Таким образом, для того чтобы провести определенную порцию воздуха через всю дыхательную систему птицы, требуются два полных дыхательных цикла. Причем оба эти дыхательных цикла абсолютно идентичны и за каждой порцией воздуха следует другая порция, повторяющая тот же путь на один цикл позже. Таким образом, наиболее примечательная особенность дыхательной системы птиц, обусловленная наличием воздушных мешков, сообщающихся с легкими, состоит в том, что воздух всегда течет через легкие по направлению от задней их области к передней и проходит через легкие как при вдохе, так и при выдохе. Такой механизм весьма выгоден для газообмена между воздухом и кровью, так как он в принципе сходен с противоточным механизмом в жабрах рыб. Благодаря ему в оксигенированной крови, покидающей легкие, парциальное напряжение кислорода может быть больше, чем в выдыхаемом воздухе. Кровь, которая должна вот-вот покинуть легкие, обменивается с воздухом, который только что вошел в легкие из задних мешков и в котором парциальное давление О2 еще не снизилось. По мере того как воздух проходит через легкие, он теряет кислород и насыщается двуокисью углерода. Но при этом на всем своем пути этот воздух встречает кровь со все более низким напряжением О2 и поэтому отдает ей все больше и больше кислорода. Благодаря этому кровь может хорошо насыщаться кислородом и способна извлекать из легочного воздуха больше кислорода и отдавать в легких больше двуокиси углерода, чем у млекопитающих. Следовательно, хотя сами по себе воздушные мешки птиц не принимают участия в газообмене с кровью, но они выполняют роль временных резервуаров воздуха и создают постоянный ток воздуха через легкие (и при вдохе, и при выдохе), что позволяет повысить эффективность насыщения крови кислородом в альвеолах легких. Эффективность однонаправленного потока воздуха в легких птиц особенно существенна на больших высотах. Так, в экспериментах на мышах и воробьях (имеют одинаковый вес и сопоставимую площадь поверхности тела), которых помещали в камеру с давлением воздуха 350 мм рт. ст. (чуть меньше 0,5 атм), соответствующим высоте 6100 м, было установлено, что мыши лежали на животе и едва могли ползать (т.е. у них развивалась тяжелая гипоксия), в то время как воробьи все еще были способны летать. Именно в связи с такой особенностью легких птиц можно объяснить их способность к полету на такой высоте в Гималаях, где альпинисты без кислородных приборов едва способны ходить. |