Биология. Итог по генетике. Генетика как наука. Предмет и задачи генетики. Наследственность и изменчивость (определение). Основные этапы развития генетики, их краткая характеристика. Роль генетики в современной биологии и медицине
 Скачать 1.66 Mb. Скачать 1.66 Mb.
|

|
Рецессивность - форма взаимоотношений двух аллельных генов, при которой один из них - рецессивный - оказывает менее сильное влияние на соответствующий признак особи, чем другой - доминантный. Рецесси́вный ген— генетическая информация, которая может подавляться воздействием доминантного гена и не проявляется в фенотипе. Рецессивный ген способен обеспечить проявление определяемого им признака только в том случае, если находится в паре с соответственным рецессивным геном. Если же он находится в паре с доминантным геном, то он не проявляется, так как доминантный ген подавляет его. Свойства, представленные рецессивными генами, проявляются в фенотипе у потомка лишь в том случае, если у обоих родителей присутствует рецессивный ген. Причина такого функционирования следующая: доминантный признак формируется аллелью, ответственной за производство определенного белка, рецессивный - аллелью, не приводящей к его производству. Достаточно наличие хотя бы одной доминантной аллели - либо в хромосоме, полученной от матери, либо в хромосоме, полученной от отца, чтобы обеспечить возможность производства этого белка и соответственно сформировать черту. Рецессивный признак определяется отсутствием формирования белка. Для того, чтобы сформировался рецессивный признак, необходимо иметь "не производящую" белок аллель в обеих хромосомах (гомозиготность) - и в хромосоме, полученной от матери, и в хромосоме, полученной от отца. В противном случае, если одна из хромосом будет содержать доминантную аллель, кодирующую белок, данный белок будет производиться организмом и сформируется доминантный признак. Сверхдоминирование – тип взаимодействия аллельных генов, при котором ген, находящийся в гетерозиготном состоянии имеет большее фенотипическое проявление признака, чем гомозиготный. Серповидно-клеточная анемия. А – гемоглобинA, а – гемоглобинS. АА – 100% нормальные эритроциты, больше подвержены малярии; аа – 100% мутированные (умирают), Аа – 50% мутированных, практически не подвержены малярии т.к. уже поражены Сверхдоминирование — более сильное проявление признака у гетерозиготной особи, чем у любой гомозиготной. На этом типе аллельного взаимодействия основано явление гетерозиса (превосходство над родителями по жизнеспособности, энергии роста, плодовитости, продуктивности). Одним из характерных примеров сверхдоминирования является повышенная частота аллеля гена серповидноклеточной анемии в популяциях человека, живущих в условиях высокой вероятности заражения малярией. Мутантный аллель защищает организм от заболевания малярией. Гомозиготы по нормальному аллелю могут заболеть малярией и погибнуть, гомозиготы по мутантному аллелю — с высокой вероятностью гибнут от анемии. Гетрозиготы по этому гену не болеют серповидноклеточной анемией и устойчивы к малярии. Преимущество гетерозигот так же показано по многим генам и у многих организмов. Для Drosophila melanogaster показаны эффекты сверхдоминирования по гену алкогольдегидрогеназы в лабораторных популяциях. В ряде случаев аллель гена, с которым связано сверхдоминирование, является рецессивно летальным и поддерживается в популяции за счёт преимущества гетерозигот. К таким случаям относится, например, система летальных аллелей гена lethal giant larvae. Гетерозиготы, имеющие нормальный и мутантный вариант этого гена, в ряде случаев, характеризуются повышенной жизнеспособностью. Пример – гетерозис, или явление гибридной силы, когда гибриды первого поколения обладают резко выраженными фенотипическими признаками (в последующих поколениях проявление этих признаков ослабевает). Гетерозис — увеличение жизнеспособности гибридов вследствие унаследования определённого набора аллелей различных генов от своих разнородных родителей. Это явление противоположно результатам инбридинга, или близкородственного скрещивания, приводящего к гомозиготности. Увеличение жизнеспособности гибридов первого поколения в результате гетерозиса связывают с переходом генов в гетерозиготное состояние, при этом рецессивные летальные и полулетальные аллели, снижающие жизнеспособность гибридов, не проявляется. Также в результате гетрозиготации могут образовываться несколько аллельных вариантов фермента, действующих в сумме более эффективно, чем поодиночке (в гомозиготном состоянии). Механизм действия гетерозиса ещё не окончательно выяснен. Явление гетерозиса зависит от степени родства между родительскими особями: чем более отдалёнными родственниками являются родительские особи, тем в большей степени проявляется эффект гетерозиса у гибридов первого поколения. Явление гетерозиса наблюдалось ещё до открытия законов Менделя И. Г. Кёльрейтером, термин «гетерозис» (в переводе с греческого языка — изменение, превращение), в 1908 Г. Шулл описал гетерозис у кукурузы У растений (по А. Густафсону) выделяют три формы гетерозиса: т. н. репродуктивный гетерозис, в результате которого повышается плодородность гибридов и урожайность, соматический гетерозис, увеличивающий линейные размеры гибридного растения и его массу, и приспособительный гетерозис (называемый также адаптивным), повышающий приспособленность гибридов к действию неблагоприятных факторов окружающей среды. Гетерозис в сельском хозяйстве. Использование Г. в растениеводстве — важный приём повышения продуктивности растений. Урожай гетерозисных гибридов на 10—30% выше, чем у обычных сортов. Для использования Г. в производстве разработаны экономически рентабельные способы получения гибридных семян кукурузы, томатов, баклажанов, перца, лука, огурцов, арбузов, тыквы, сахарной свёклы, сорго, ржи, люцерны и др. с.-х. растений. Особое положение занимает группа вегетативно размножаемых растений, у которых возможно закрепление Г. в потомстве, например сорта картофеля и плодово-ягодных культур, выведенные из гибридных семян. Для использования Г. с практической целью применяются межсортовые скрещивания гомозиготных сортов самоопыляющихся растений, межсортовые (межпопуляционные) скрещивания самоопылённых линий перекрёстноопыляющихся растений (парные, трёхлинейные, двойные — четырёхлинейные, множественные) и сортолинейные скрещивания. Преимущество определённых типов скрещивания для каждой с.-х. культуры устанавливается на основе экономической оценки. Устранению трудностей В получении гибридных семян может способствовать использование цитоплазматической мужской стерильности (ЦМС), свойства несовместимости у некоторых перекрестноопыляющихся растений и других наследственных особенностей в структуре цветка и соцветия, исключающих большие затраты на кастрацию. При выборе родительских форм для получения гетерозисных гибридов оценивают их комбинационную способность. Первоначально селекция в этом направлении сводилась к выделению лучших по комбинационной ценности генотипов из популяций свободноопыляющихся сортов на основе инбридинга в форме принудительного самоопыления. Разработаны методы оценки и повышения комбинационной способности линий и др. групп растений, используемых для скрещиваний. Наибольший эффект в использовании Г. достигнут на кукурузе. Создание и внедрение в производство гибридов кукурузы позволило повысить на 20—30% валовые сборы зерна на огромных площадях, занимаемых этой культурой в разных странах мира. Созданы гибриды кукурузы, совмещающие в себе высокую урожайность с хорошим качеством семян, засухоустойчивостью и иммунитетом к различным болезням. Районированы гетерозисные гибриды сорго (Гибрид Ранний 1, Гибрид Восход), гетерозисные межсортовые гибриды сахарной свёклы, из которых наибольшее распространение получил Ялтушковский гибрид. Для получения гетерозисных форм все шире используются линии сахарной свеклы со стерильной пыльцой. Явления Г. установлены также у многих овощных и масличных культур. Получены первые результаты в изучении Г. у гибридов пшеницы первого поколения, созданы стерильные аналоги и восстановители фертильности (плодовитости), выявлены источники ЦМС у пшеницы. В животноводстве явления Г. наблюдаются при гибридизации, межпородном и внутрипородном (межлинейном) скрещивании и обеспечивают заметное повышение продуктивности с.-х. животных. Наибольшее распространение получило использование Г. при промышленном скрещивании. В птицеводстве при скрещивании яйценоских пород кур, например леггорнов с австралорпами, родайландами и др., яйценоскость помесей первого поколения возрастает на 20—25 яиц в год; скрещивание мясных пород кур с мясо-яичными обусловливает повышение мясных качеств (см. Бройлер); Г. по комплексу признаков получают при скрещивании близкородственных линий кур одной породы или при межпородных скрещиваниях. В свиноводстве, овцеводстве и скотоводстве промышленным скрещиванием пользуются для получения Г. по мясной продуктивности, что выражается в повышении скороспелости и живого веса животных, увеличении убойного выхода, улучшении качества туши. Свиней мясо-сальных (комбинированных) пород скрещивают с хряками мясных пород. Мелких малопродуктивных овец местных пород скрещивают с баранами мясо-шёрстных пород, тонкорунных маток — с баранами скороспелых мясных или полутонкорунных пород. Для повышения мясной продуктивности коров молочных, молочно-мясных и местных мясных пород скрещивают с быками специализированных мясных пород. Кодоминирование – проявление в гетерозиготном состоянии признаков, кодируемых обоими аллельными генами. Примеры: гены нормального и серповидноклеточного гемоглобина; наследование у человека IV группы крови (AB). В то же время группы крови являются примером множественного аллелизма. Множественный аллелизм – наличие в генофонде популяции более двух аллельных генов. Пример. Окраска шерсти у кроликов определяется четырьмя аллельными генами: A, ach, ah, a. A – ген, определяющий черную окраску (дикий тип); ach – ген шиншилловой окраски; ah – ген гималайской окраски; a – ген белой окраски. Характер их взаимодействия: A > ach > ah > a. Группы крови человека по системе АВО кодируются тремя аллельными генами: IA, IB, I0. Группа крови Генотип 0 (I) I0 I0 А (II) IA I0, IA IA; B (III) IB I0, IB IB; AB (IV) IA IB (фенотипически проявляется действие обоих аллельных генов – явление кодоминирования). Иногда к числу аллельных может относиться не два, а большее число генов. Они получили название множественных аллелей. Возникают множественные аллели в результате многократного мутирования одного и того же локуса в хромосоме. Так, кроме основных доминантного и рецессивного аллельных генов, между ними возникают промежуточные, которые по отношению к доминантному ведут себя как рецессивные, а по отношению к рецессивному - как доминантные гены. Генетико-физиологическая характеристика системы АВ0 С точки зрения генетики, наиболее изученной является система АВ0, определяющая I (0), II (А), III (В) и IV (АВ) группы крови. На поверхности эритроцитов могут находиться агглютиногены (антигены) А и В, а в плазме крови – агглютинины (антитела) и . В норме одноименные агглютиногены и агглютинины совместно не обнаруживаются. Нужно отметить, что А- и В-антигены образуют многочисленный ряд антигенов (А1, А2 ... A; В1, В2 …В). Понятие о неаллельных генах. Межаллельная комплементация и аллельное исключение (примеры, типы расщепления). Эпистатическое взимодейстивие неаллельных генов (примеры, типы расщепления). Неаллельные гены - это гены, которые расположены в негомологичных хромосомах. Взаимодействие между неаллельными генами происходит в том случае, если признак проявляется только при их совместном присутствии. Межаллельная комплементация – вид взаимодействия аллельных генов, при котором возможно формирование нормального признака у организма, гетерозиготного по двум мутантным аллелям этого гена. Пример: D – ген, кодирующий синтез белка с четвертичной структурой (например, глобин в гемоглобине). Четвертичная структура состоит из нескольких полипептидных цепей. Мутантные гены – Dи D – определяют синтез измененных белков (каждый своего). Однако при объединении этих цепей в четвертичной структуре образуется белок с нормальными свойствами: D + D = D. К сравнительно редким формам взаимодействия аллельных генов относятся межаллельная комплементация (взаимодополнение) и аллельное исключение. О межаллельной комплементации говорят тогда, когда у организма, гетерозиготного по двум мутантным аллелям конкретного гена, в фенотипе обнаруживается признак в нормальном, то есть наиболее часто встречаемом состоянии («дикий тип»). Допустим, что ген D контролирует образование клеткой белка с четвертичной структурой в виде комплекса из нескольких одинаковых полипептидов — мультигомобелковый комплекс. Один мутантный аллель D́ определяет экспрессию измененного полипептида D ́, а второй аллель D´´ — тоже мутантный, но по другому участку гена, определяет экспрессию мутантного полипептида D΄΄, причем с другим изменением аминокислотной последовательности. Допускается, что при формировании четвертичной структуры с участием измененных, но по-разному, полипептидов D´ и D́́΄ происходит компенсация изменений и, в итоге, формируется сложный мультигомобелковый комплекс с нормальной функцией. Суть аллельного исключения поясняет пример генетической инактивации одной из хромосом Х у особей гомогаметного пола (у человека — женский, 46ХХ), функционально-генетический смысл которой заключается в компенсации дозы генов соответствующей группы сцепления (хромосомы Х) относительно гетерогаметного пола (у человека — мужской, 46ХУ). По хромосоме Х гомогаметный пол представлен особями-«мозаиками» — одна хромосома Х материнского, тогда как другая отцовского происхождения. Так как генетическая инактивация носит относительно хромосом Х материнского и отцовского происхождения случайный характер, при гетерозиготности организма в одних клетках активен аллель, полученный с хромосомой Х матери, тогда как в других активен аллель, полученный с хромосомой Х отца. Это приводит к фенотипическому мозаицизму (рис. 4-4). 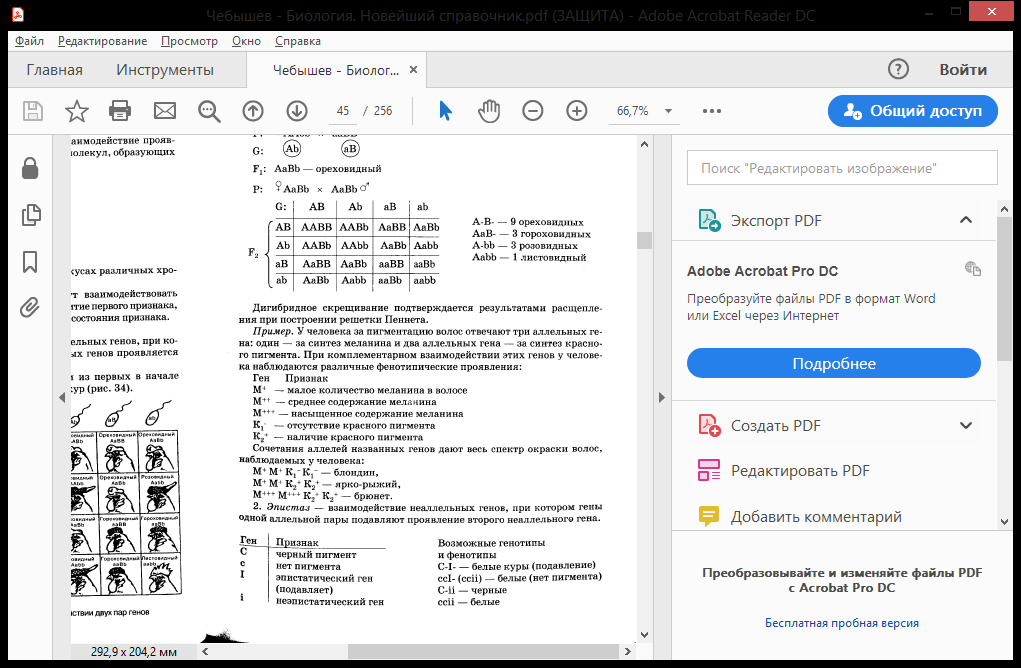 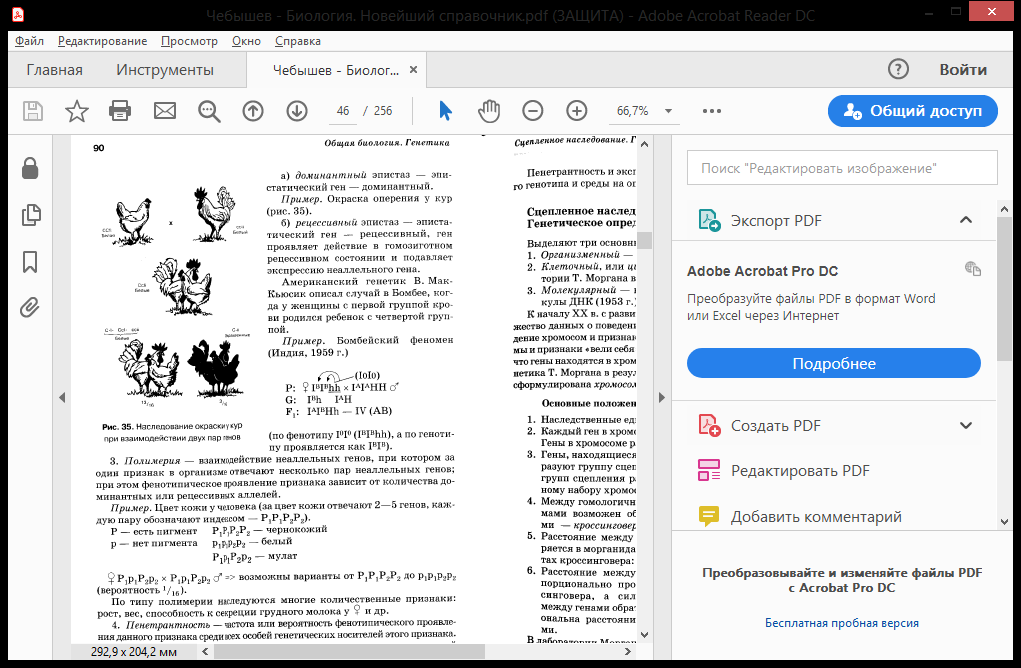 Эпистаз – такая форма взаимодействия неаллельных генов между собой, при которой гены одной аллельной пары подавляют действие генов другой аллельной пары. Ген, подавляющий действие другого гена называется эпистатическим геном (супрессором), а подавляемый им ген носит название гипостатического. Выделяют доминантный и рецессивный эпистаз. Доминантный эпистаз – эпистатическое действие оказывает доминантный ген в гомо- или гетерозиготном состоянии (например, окраска оперения у кур). Рецессивный эпистаз – эпистатическое действие оказывает рецессивный ген только в гомозиготном состоянии (например, окраска шерсти у домовой мыши). При доминантном эпистазе при скрещивании двух гомозиготных особей, отличающихся по двум парам неаллельных генов, расщепление по фенотипу среди гибридов второго поколения будет выражаться соотношением 12:3:1 или 13:3. При рецессивном эпистазе это соотношение 9:3:4. Примером доминантного эпистаза может служить наследование окраски оперения у кур. У большинства пород кур (плимутрок, виандот) доминантный ген С обусловливает развитие пигментированного оперения, а его аллель с — белого (альбинизм). У породы леггорн существует эпистатический ген I, подавляющий действие доминантного гена С. Его аллель — i не влияет на проявление гена С. Леггорны с генотипом IICC имеют белое оперение в связи с эпистазом гена I над геном С. Виандоты с генотипом iicc имеют белое оперение вследствие отсутствия доминантного аллеля пигментации — С. Проведено скрещивание гомозиготного белого леггорна (IICC) с белым виандотом (iicc). Потомство от этого скрещивания все оказалось белым (IiCc). Далее при скрещивании гибридов первого поколения между собой в F2 наблюдалось следующее расщепление: 13/16 белых кур (I-C-, Icc, iicc) и 3/16 кур с пигментированным оперением (iiC-). В целом насле- дование окраски оперения у кур, иллюстрирующее доминантный эпистаз можно представить в схеме 7. Другими примерами доминантного эпистаза могут являться наследование масти у лошадей, окраски плодов тыквы, окраски тела у свиней. Явление рецессивного эпистаза можно проиллюстрировать на приме- ре наследования пигментации шерсти у мышей. Доминантный ген А определяет зональное распределение пигмента по длине волоса у мышей, а его аллель а не влияет на зональное распределение пигмента. Доминантный ген С детерминирует синтез пигмента, а его аллель с определяет отсутствие пигмента (альбинизм) и не дает проявиться гену А (а) т.е. является эпистатическим геном. Произведено скрещивание гомозиготной черной особи (CCаа) с гомо- зиготной белой особью (ссАА). Потомство от этого скрещивания оказалось серым (СсАа). Далее при скрещивании гибридов первого поколения между собой в F2 отмечено следующее расщепление: 9/16 серых мышей (С-А-), 3/16 черных (С-аа) и 4/16 белых (ссА-, ссаа). В целом наследование пигментации шерсти у мышей, демонстрирующее рецессивный эпистаз можно представить в схеме 8. Примерами рецессивного эпистаза у человека могут являться случаи альбинизма и «бомбейский феномен». «Бомбейский феномен» впервые описан в семье, где у женщины с I группой крови родился ребенок с IV группой крови. Он связан с тем, что в формировании группы крови системы АВО наряду с аллелями гена I (I0 , IА, IВ ), определяющими синтез анти- генов А и В, задействован и ген Н, детерминирующий синтез вещества- предшественника из которого в последующем образуются антигены А и В. У гомозигот по рецессивному аллелю h (hh) даже при наличии в генотипе доминантных аллелей гена I (IА и IВ ) антигены А и В не синтезируются, и фенотипически эти люди имеют I группу крови. Варианты расщепления в F2 : 12:3:1, 13:3. |