Биология. Итог по генетике. Генетика как наука. Предмет и задачи генетики. Наследственность и изменчивость (определение). Основные этапы развития генетики, их краткая характеристика. Роль генетики в современной биологии и медицине
 Скачать 1.66 Mb. Скачать 1.66 Mb.
|

|
Генетический пол. Различия в формировании мужского и женского организма обнаруживаются с первых недель его развития. Все клетки человека, за исключением половых, имеют 23 пары хромосом, из которых одна пара — половые. Половые клетки (гаметы) — яйцеклетки и сперматозоиды — образуются путем особого деления — мейоза, отличающегося от обычного деления — митоза — тем, что при этом вновь возникшие клетки получают одну (сромосому из каждой пары. Генетический пол определяется отцовской половой клеткой (спермато-|зоидом) во время оплодотворения. Это связано с тем, что развитие женско-i пола у человека обусловлено наличием в зиготе (оплодотворенной яйцеклетке) двух половых хромосом X и X, а мужского — X и Y. Все яйцеклетки в результате мейоза имеют одинаковые хромосомы — X, тогда как спермато- 380 381 зо 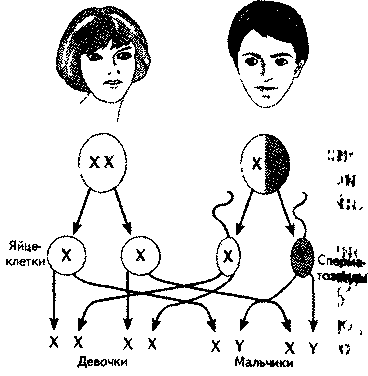 иды могут быть двух типов: сX-или с Y- хромосомами. В зависимости от того, какой сперматозоид оплодотворит яйцеклетку, зародыш будет либо мужским, либо женским (рис. 17.1). иды могут быть двух типов: сX-или с Y- хромосомами. В зависимости от того, какой сперматозоид оплодотворит яйцеклетку, зародыш будет либо мужским, либо женским (рис. 17.1).Частота Первичная детерминация пола. Схема определения пола у млекопитающих может выглядеть так. На догонадной стадии у эмбриона отсутствуют половые органы. На прогонадной — выделяются гонадный валик и бисексуальные гонады, предшественники и мужских, и женских половых органов. Предшественником женских половых органов является Мюллеро-ва система, мужских — Вольфова система. Мюллерова система в дальнейшем дает начало фаллопиевым трубам, матке, влагалищу. Из Воль-фовой системы развиваются яички, придатки яичек, семявыносящий проток, предстательная железа (Carlson, 1992). Рис. 17.2. Развитие внутренних половых ор-| ганов (Carlson, 1992). Первичная закладка пола связана с программированием Y-хромосомой синтеза фактора, определяющего развитие яичек. Дифференциров-ка гонад у человеческого зародыша происходит приблизительно на шестой неделе развития, когда у XY-зародыша образуются яички, а у \Рис 173. Развитие внешних половых органов ((Carlson, 1992) Х 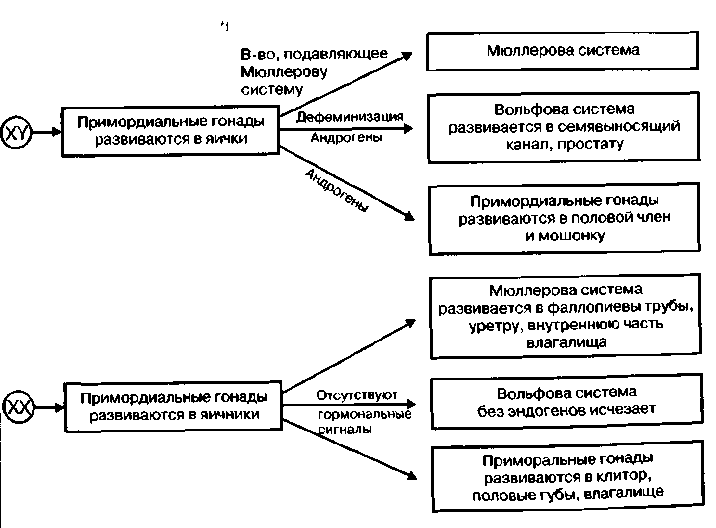 Х-зародыша — яичники (рис. 17.2). Это приводит к появлению клеток Сертоли, окружающих семявыносящие канальцы. Клетки Сертоли, в свою очередь, синтезируют фактор, подавляющий развитие Мюллеровых протоков, что ведет к формированию Вольфовых протоков (рис. 17.2). Позднее в них возникают клетки Лейдига, синтезирующие тестостерон и дигидро-тестостерон, которые стимулируют развитие Вольфовой системы (Смирнов, 1977). Х-зародыша — яичники (рис. 17.2). Это приводит к появлению клеток Сертоли, окружающих семявыносящие канальцы. Клетки Сертоли, в свою очередь, синтезируют фактор, подавляющий развитие Мюллеровых протоков, что ведет к формированию Вольфовых протоков (рис. 17.2). Позднее в них возникают клетки Лейдига, синтезирующие тестостерон и дигидро-тестостерон, которые стимулируют развитие Вольфовой системы (Смирнов, 1977). Синтез тестис-определяющего фактора связывают с активностью гена SRY (sex determinging region Ygene), открытого в 1976 г. (Berta e. а., 1992; Koopman e. а., 1992). Этот ген у человека имеет небольшой размер, не содержит интронов (вставок, непосредственно не определяющих структуру белка), кодирует белок размером в 204 аминокислотных остатка. Это лишь один ген из большого семейства (их около 20) (Смирнов, 1997). Поскольку эмбрионы и гонады будущих самцов растут быстрее, чем самок, еще до развития гонадного бугорка можно предположить, что есть и другие факторы, предопределяющие дифференциацию прогонад по мужскому или женскому типу. Был найден ген DAX1, кодирующий ядерный рецептор. При дупликации (удвоении) он может вызывать возвращение мужского пола к женскому. Он чувствителен к гормонам и при высокой их концентрации способен преодолеть сигнал SRY гена и сдвинуть развитие прогонад в направлении яичника. Этот ген рассматривается как реликт более примитивной Х-хромосом-ной системы детерминации пола. Предполагается, что активность SRY гена является недостаточным усло вием появления семенников из потенциально бисексуальных гонад. Для появления клеток Сертоли, семявыносящих канальцев, клеток Лейдига и т. д. требуется его взаимодействие со многими аутосомальными и Х-хромосом- ными факторами. В эволюционном развитии все более усиливается взаимо связь гаметного и соматического полов. Направление дифференциации по ловых клеток на каждом этапе развития зависит не только от их собствен ной хромосомной конституции, но и от окружения, созданного соматичес кими клетками гонад. Детерминация пола в этом случае является сложным многокомпонентным процессом (Смирнов, 1997). , В отличие от Вольфовой системы Мюллерова не нуждается в гормональной стимуляции, поэтому при всех сбоях в последовательности активации генов, кодирующих факторы дифференцировки гонад, формируется женский зародыш. Мюллерова система имеет рецепторы к гормону, подавляющему ее собственное развитие, а Вольфова чувствительна только к андро-генам. Активность андрогенов продолжается приблизительно до 32-й недели, а затем клетки Лейдига претерпевают обратное развитие. Формирование женских и мужских половых органов. Уже отмечалось, что синдром Тернера связан с наличием только одной Х-хромосомы (ХО) у человека, когда отсутствует и вторая Х-хромосома и Y-хромосома. У таких организмов нет ни мужских, ни женских гонад, но развиваются женские внутренние и наружные половые органы, поскольку для этого не нужны до- 3 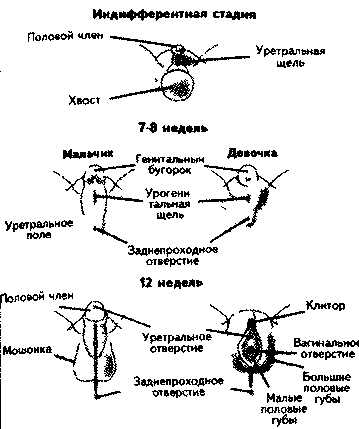 84 84полнительные факторы. Внутренние половые органы женщины формируются из Мюллеровой системы, а наружные — под воздействием гормонов, вырабатываемых фетальными (fetus — плод, лат.) женскими половыми органами. Развитие внешних половых органов мужчины Определяется активностью яичек и выработкой ими фетальных андрогенов (рис. 17.3). Факторы, участвующие в становлении внутренних и внешних половых органов, обозначены на рис. 17.4. С активностью фетальных гормонов связана и дифференцировка определенных отделов мозга, регулирующих половые различия в поведении. Половые хромосомы (гоносомы, гетеросомы) различаются как по строению (длина, положение центромеры, количество гетерохроматина), так и по содержанию генов. Хромосома X - это субметацентрическая хромосома средних размеров, входит в группу С). Она есть в соматических клетках индивидов обоих полов: в двойном экземпляре у женщин с кариотипом 46,ХХ и в одном экземпляре у мужчин с кариотипом 46,ХY; а также в одном экземпляре во всех яйцеклетках и 50% сперматозоидов.Хромосома X богата эухроматиновыми участками и содержит 1336 генов, среди которых: соматические гены, регуляторные гены феминизации, структурные гены феминизации, структурные гены маскулинизации. Таким образом, хромосома Х является обязательной в кариотипе соматической клетки как женского, так и мужского полов. Хромосома Y является мелкой акроцентрической хромосомой, входит в группу G; 2/3 дистального плеча q представлены гетерохроматином и генетически неактивны. Хромосома Y представлена одним экземпляром во всех соматических клетках индивидов мужского пола с кариотипом 46,XY и у 50% сперматозоидов. Она содержит 307 генов, среди которых: регуляторные гены маскулинизации (SRY + Tdf), гены, обеспечивающие фертильность (AZF1, AZF2), несколько структурных соматических генов и псевдогены. Морфологические и генетические различия между хромосомами X и Y, а также отличия по количеству хромосом Х в кариотипе стало причиной генетического неравенства между полами (у женщин по сравнению с мужчинами двойная доза генов хромосомы Х. Однако это неравенство не проявляется, благодаря механизму компенсации, в результате которого функциональной остается только одна хромосома Х в соматических клетках и мужчин и женщин, а именно: - в клетках 46,ХХ – активна только одна хромосома Х; - в клетках 46,XY – активны хромосомы Х и Y; - в клетках 47,ХХХ – активна только одна хромосома Х; - в клетках 47,ХХY – активна только одна хромосома Х и одна хромосома Y; - в клетках 48,ХХХY – активна только одна хромосома Х и одна Y; Путем гетерохроматинизации одной из двух хромосом Х и женщин образуется половой хроматин Х, а в результате гетерохроматинизации 2/3q хромосомы Y у лиц мужского пола образуется половой хроматин Y. Половой хроматин Х: - представляет инактивированную хромосому Х. в форме факультативного гетерохроматина, в соматических клетках 46,ХХ; - выявляется в интерфазных ядрах соматических клеток в виде тельца Барра размером около 1µm; - тест Барра используется для определения количества хромосом Х в кариотипе в норме и в случае гносомных анеуплоидий; - число хр.Х = числу телец Барра + 1 (активная хр.Х); 46,ХХ – 1 тельце Барра; 46,ХY – отсутствует тельце Барра; 47,ХХХ – 2 тельца Барра; 47,ХХY – 1 тельце Барра; 45,Х – отсутствует тельце Барра; 48,ХХХХ – 3 тельца Барра. Половой хроматин Y: - представлен 2/3 плеча Y q хромосомы Y, в форме конститутивного гетерохроматина, в соматических клетках 46,XY и сперматозоидах 23,Y; - выявляется в интерфазных ядрах клеток в виде тельца F (флуоресцентного) размером около 1µm; - тест F используется для идентификации хромосомы Y (пренатальное определение пола); - число хр.Y = числу телец F; 46,ХХ – отсутствует тельце F; 46,ХY – 1 тельце F; 47,ХYY – 2 тельца F; 47,ХХY – 1 тельце F; 48,ХХYY – 2 тельца F; 46,X,i(Yp) – отсутствует тельце F; 46,X,i(Yq) – 1(0,5 µm) тельце F. В 1949 г. М.Барр и Ч.Бертрам обнаружили в ядрах нервных клеток кошки крупную глыбку хроматина. Она выявлялась у самок и отсутствовала у самцов. Позже было установлено, что это спирализованная Х-хромосома. Эту глыбку хроматина назвали тельцем Барра. Тельце Барра может быть прикреплено к ядрышку, к ядерной оболочке, В 1962 году Мари Лайон высказала гипотезу об инактивации одной Х-хромосомы у женских особей млекопитающих.Каждая клетка женского зародыша содержит две Х-хромосомы – одну материнскую (Хм), вторую – отцовскую (Хо). До 16-го дня эмбриогенеза при дроблении в каждую клетку попадают две активные Х-хромосомы. На 16-й день происходит инактивация одной из Х-хромосом – с одинаковой вероятностью – отцовской или материнской. Процесс инактивации случайный,поэтому в половине клеток остается активной отцовская Х-хромосома, в другой половине клеток активной будет материнская Х-хромосома. Материнская и отцовская Х-хромосомы содержат аллельные гены (доминантные и рецессивные). В клетках могут синтезироваться два вариантаферментов, которые различаются по сродству к субстрату или по рН-показателям. Это обеспечивает женским организмам более широкие возможности адаптаций к условиям окружающей среды. Суть женского мозаицизма по половым хромосомам – содержание в разных клетках разных активных Х-хромосом – от отца и от матери. Теоретически соотношение мужских и женских гамет (соотношение полов) в момент оплодотворения составляет примерно 1 : 1. Первичное соотношение полов в момент зачатия: на 100 женских зигот 140-150 мужских зигот.Объяснить это можно тем, что сперматозоиды с Y-хромосомой легче, подвижнее, имеют отрицательный заряд (у яйцеклетки он положительный) и обладают положительным хемотаксисом на вещества, выделяемые яйцеклеткой. Сперматозоиды, содержащие Y-хромосому, чаще оплодотворяют яйцеклетку. Вторичное соотношение полов (в момент рождения): 100 ♀ : 106 ♂ Такое соотношение можно объяснить большей жизнестойкостью женских зигот (по гипотезе М.Лайон), гемизиготностью мужских зигот (у них проявляются все рецессивные гены) и чужеродностью (по белкам) для материнского организма мужской зиготы. Вторичное соотношение полов может зависеть от ряда факторов: 1) от возраста матери (при беременности) - при возрасте 18-20 лет на 100♀ - 120♂; при возрасте 32-40 лет на 100♀ - 90♂; 2) при первых родах чаще бывают мальчики; 3) при токсикозах беременности и стрессовых воздействиях – больше рождается девочек. Третичное соотношение (постнатальный период): к 20-ти годам – на 100 ♀ : 100 ♂; к 50-ти годам – на 100 ♀ : 85 ♂; к 80-ти годам – на 100 ♀ : 50 ♂. Это соотношение также можно объяснить большей жизнестойкостью женского организма и большей гибелью мужчин в постнатальный период (от болезней, в войнах, при выполнении тяжелого физического труда, от вредных привычек, в автокатастрофах). Анализ наследования признака окраски глаз у дрозофилы в лаборатории Т. Моргана выявил некоторые особенности, заставившие выделить в качестве отдельного типа наследования признаков сцепленное с полом наследование. Зависимость результатов эксперимента от того, кто из родителей являлся носителем доминантного варианта признака, позволила высказать предположение, что ген, определяющий окраску глаз у дрозофилы, расположен в Х-хромосоме и не имеет гомолога в Y-хромосоме. Все особенности сцепленного с полом наследования объясняются неодинаковой дозой соответствующих генов у представителей разного— гомо- и гетерогаметного пола. Гомогаметный пол несет двойную дозу генов, расположенных в Х-хромосоме. Развитие соответствующих признаков у гетерозигот (ХAХa) зависит от характера взаимодействия между аллельными генами. Гетерогаметный пол имеет одну Х-хромосому (ХО или XY). У некоторых видов Y-хромосома генетически инертна, у других она содержит некоторое количество структурных генов, часть из которых гомологична генам Х-хромосомы. Гены негомологичных участков Х- и Y-хромосом (или единственной Х-хромосомы) у гетерогаметного пола находятся вгемизиготномсостоянии. Они представлены единственной дозой: ХAY, ХaХ, XYB. Формирование таких признаков у гетерогаметного пола определяется тем, какой аллель данного гена присутствует в генотипе организма. Характер наследования сцепленных с полом признаков в ряду поколений зависит от того, в какой хромосоме находится соответствующий ген. В связи с этим различают Х-сцепленное и Y-сцепленное (голандрическое) наследование. Х-сцепленное наследование. Х-хромосома присутствует в кариотипе каждой особи, поэтому признаки, определяемые генами этой хромосомы, формируются у представителей как женского, так и мужского пола. Особи гомогаметного пола получают эти гены от обоих родителей и через свои гаметы передают их всем потомкам. Представители гетерогаметного пола получают единственную Х- хромосому от гомогаметного родителя и передают ее своему гомогаметному потомству. У млекопитающих (в том числе и человека) мужской пол получает Х- сцепленные гены от матери и передает их дочерям. При этом мужской пол никогда не наследует отцовского Х-сцепленного признака и не передает его своим сыновьям. Так как у гомогаметного пола признак развивается в результате взаимодействия аллельных генов, различают Х-сцепленное доминантное и Х-сцепленное рецессивное наследование. Х-сцепленный доминантный признак(красный цвет глаз у дрозофилы) передается самкой всему потомству. Самец передает свой Х-сцепленный доминантный признак лишь самкам следующего поколения. Самки могут наследовать такой признак от обоих родителей, а самцы — только от матери. Х-сцепленный рецессивный признак, (белый цвет глаз у дрозофилы) у самок проявляется только при получении ими соответствующего аллеля от обоих родителей (XaXa). У самцов XaY он развивается при получении рецессивного аллеля от матери. Рецессивные самки передают рецессивный аллель потомкам любого пола, а рецессивные самцы — только «дочерям». При Х-сцепленном наследовании, так же как и при аутосомном, возможен промежуточный характер проявления признака у гетерозигот. Например, у кошек пигментация шерсти контролируется Х-сцепленным геном, разные аллели которого определяют черную (XAи рыжую (XA’) пигментацию. Гетерозиготные самки XAXA’ имеют пеструю окраску шерсти. Самцы же могут быть либо черными (XAY, либо рыжими (XA’Y). Голандрическое наследование. Активно функционирующие гены Y-хромосомы, не имеющие аллелей в Х-хромосоме, присутствуют в генотипе только гетерогаметного пола, причем в гемизиготном состоянии. Поэтому они проявляются фенотипически и передаются из поколения в поколение лишь у представителей гетерогаметного пола. Так, у человека признак гипертрихоза ушной раковины («волосатые уши») наблюдается исключительно у мужчин и наследуется от отца к сыну. У человека X-хромосома, которую мужчина получает от матери, несет геныдальтонизма(цветовой слепоты) игемофилии(несвертываемости крови). Эти гены рецессивны; у женщин названные болезни проявляются крайне редко, а у мужчин чаще, так как в Y-xpoмосомах мужчин нет доминантного аллеля, подавляющего действие этих генов,Y-хромосоманесет такие признаки, как, на пример,волосатость мочки ушей,перепонка между пальцами ног, поэтому эти признаки проявляются только у мужчин как носителей Y-хромосомы. Всего с полом сцеплено около 120 признаков. Признаки голандрические - признаки, которые детерминируются генами негомологичного участка Y-хромосомы. В отличие от аутосом и Х-хромосомы, Y-хромосома несет сравнительно мало генов (по последним данным международного каталога генов OMIM, всего около 40). Небольшая часть таких генов гомологична генам X-хромосомы, остальные присутствующие только у мужчин, участвуют в контроле детерминации пола и сперматогенеза. Так, на Y-хромосоме находятся гены SRY и AZF, ответственные за программу половой дифференцировки. Мутации в любом из этих генов приводят к нарушениям развития яичек и блоку сперматогенеза, что выражается в азооспермии. Такие мужчины страдают бесплодием, и потому их заболевание не наследуется. Мужчин с жалобами на бесплодие необходимо обследовать на наличие мутаций в указанных генах. Мутациями в одном из генов, расположенных на Y-хромосоме, обусловлены некоторые формы ихтиоза (рыбья кожа). Примеры признаков: гипертрихоз ушных раковин, избыточный рост волос на средних фалангах пальцев кистей, азооспермия, синдактилия, повышенное ороговение кожи, ихтиоз. Особенности: передача признака от отца всем сыновьям и только сыновьям, дочери никогда не наследуют признак от отца, больные во всех поколениях, вертикальный характер наследования признака, вероятность наследования для лиц мужского пола равна 100%. |