Курс лекций по общей микробиологии основам вирусологии двух частях
 Скачать 4.07 Mb. Скачать 4.07 Mb.
|

|
6.2. Функциональная структура сапрофитной группировки почвенных микроорганизмов Микробная система почв включает различные функциональные группы (рис. 25). Взаимосвязь между ними осуществляется на основе по-следовательного потребления ресурсов питания и биохимических меха-низмов регуляции процессов разложения органических веществ. Все бак-терии и грибы - осмотрофные организмы и не способны потреблять нерас-творимые полимерные соединения типа крахмала, клетчатки или лигнина до тех пор, пока они не будут разложены до соответствующих раствори-мых мономеров. Процесс гидролиза полимеров осуществляют микроорга-низмы, составляющие группу гидролитиков, которые выделяют в среду внеклеточные гидролитические ферменты. К гидролитикам относят грибы и многие грамположительные бактерии, включая актиномицеты. Время активного роста гидроли-тиков в почве обычно непродолжительно, так как накопление мономеров тормозит их рост по типу катаболит-ной репрессии. Вспышка роста гидролитиков сменяется переходом к покою, что обеспечивается их 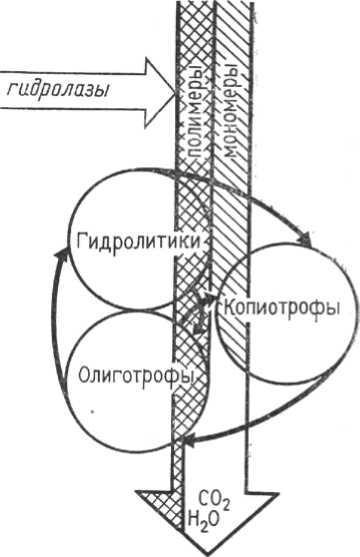 способностью образовывать споры. Выделенные в среду ферменты могут продолжать участвовать в процессах гидролиза полимеров. Таким образом, скорость превращения веществ в почве определяется концентрацией ферментов - гидро-лаз, продуцируемых микроорганизмами группы гидролитиков. Накопление мономерных соеди- Рис. 25. Схема функционирования нений типа простых углеводов и ами-гетеротрофной части микробной нокислот приводит к активному росту системы почв [12] микроорганизмов копиотрофов, которые не образуют гидролаз, но с большой скоростью потребляют растворимые соединения при их высокой концентрации в среде. В эту группу входят «сахаролитические» грибы, дрож-158 жи и многие грамотрицательные бактерии. По мере снижения концентра-ции мономеров рост копиотрофов замедляется и на смену им приходят микроорганизмы олиготрофы. Они имеют систему транспорта мономер-ных соединений в клетку в условиях низкой концентрации их в среде. Олиготрофная группа микроорганизмов представлена в почвах в основном неспорообразующими грамотрицательными бактериями, часто необычной морфологии: многие из них имеют различные придатки, образуют вырос-ты, простеки. В отличие от гидролитиков и копиотрофов они длительно активны в почве и составляют группу основных утилизаторов органиче-ских веществ на конечных стадиях их превращения. Таким образом, микробная система почвы, обеспечивающая минера-лизацию органических веществ, может быть представлена в виде триады, составленной последовательно функционирующими группами гидролити-ков, копиотрофов и олиготрофов. Регуляция активности каждой из этих групп основана на изменении концентрации мономерных субстратов (принцип катаболитной репрессии, субстратного ингибирования) и обес-печивается благодаря механизмам обратной связи. 6.3. Анализ физиологических групп бактерий Анализ физиологических групп дает возможность составить пред-ставление о соотношении микроорганизмов, осуществляющих различные физиологические процессы, и до некоторой степени судить о господ-ствующих направлениях в этих процессах. каждую физиологическую группу обычно входят совершенно раз-ные в систематическом отношении микроорганизмы, но они объединяются общностью осуществляемых ими превращений. Наиболее показательным для оценки почвенного плодородия счита-ют групповой состав микроорганизмов, связанных с циклом превращения азота, а также целлюлозоразлагающих микроорганизмов. Устанавливаются определенные закономерности в распределении разных групп микроорга-низмов в почвах разных типов. Например, в бедных питательными вещест-вами кислых подзолистых почвах обычно слабо выражены или совсем от-сутствуют нитрификаторы и азотобактер. Наличие большого количества микроорганизмов, участвующих в процессах азотфиксации, нитрифика-ции, разрушении целлюлозы, говорит о плодородии почв. Следует отме - 159 тить, что большое количество денитрификаторов не может рассматривать-ся как отрицательное явление. Денитрификация осуществляется ими толь-ко в определенных условиях (отсутствие кислорода воздуха и наличие большого количества окисленных форм азота). В других условиях они мо-гут оказывать благоприятное действие на развитие растений, так как обра-зуют полезные физиологически активные вещества. Изучение интенсивности разложения целлюлозы дает возможность судить о скорости распада растительных остатков, в состав которых всегда входит значительное количество клетчатки, а также до некоторой степени об обеспеченности почвы азотом, так как интенсивное протекание этого процесса невозможно без достаточного количества связанных форм азота. зависимости от типа питания бактерии делятся на гетеротрофные и автотрофные. Гетеротрофные бактерии осуществляют один из главнейших для ка-ждой почвы процессов - разложение органических остатков до простых минеральных соединений, которое происходит в зависимости от характера воздушного режима в аэробных или анаэробных условиях. Аэробные бак-терии осуществляют окисление белков, жиров, углеводов и других слож-ных органических соединений, являющихся компонентами мертвых расти-тельных и микробных остатков, до аммиака, воды и углекислого газа. Сре-ди аэробных гетеротрофных бактерий в почве широко распространены как спорообразующие (Bac.mycoides, Bac.subtilis и др.), так и не образующие спор (P seudomonas fluorescens, Pseudomonaspyacyanea и др.). Анаэробные бактерии вызывают процессы гниения компонентов растительных и микробных клеток также до простых, но недоокисленных органических, а затем минеральных соединений. Для создания благопри-ятных условий питания зеленых растений существенным является процесс аммонификации - разложения белковых соединений до аммиака: белок -^ аминокислоты -^ NH3 + органические безазотистые соединения. Аммонификация осуществляется как аэробными, так и анаэробными гетеротрофами. Различают окислительную, гидролитическую, восстанови-тельную аммонификации, в результате которых в почве наряду с аммиаком образуются различные органические кислоты и спирты. Анаэробные гете-ротрофные бактерии вызывают различные типы брожений углеводов, а также процессы денитрификации и десульфофикации. 160 Наиболее широко в почвах распространено масляно-кислое броже-ние различных углеводов (клетчатки, гемицеллюлоз, пектиновых веществ), при котором происходит расщепление углеводов на масляную кислоту, уг-лекислоту и водород по суммарному уравнению: 6H12 O6 ■* CH3 CH2 CH2 COOH + 2CO2 + 2H2 + 193 кДж. Этот процесс осуществляют различные виды Clostridium. Пектино-вые вещества сбраживает Clostridium pektinovorum. Главным представите-лем анаэробных целлюлозоразлагающих бактерий является Clostridium omelianski. При высоких температурах разложение клетчатки вызываютспециальные термофильные бактерии Clostridium thermocellum. Денитрификация - восстановление нитратов до молекулярного азо-та, осуществляется группой факультативных анаэробных гетеротрофов, использующих для питания углеводы (глюкозу): 5C6 Ни О6 + 24KNO3 ■* 24 KHCO3 + 6CO2 + 12N2 + 18H2 O + 1722 кДж. Основными денитрификаторами в почве являются P seudomonas fluo-rescens, Paracoccus denitrificans и др.Этот процесс приводит к значитель-ным потерям азота в почвах с плохой аэрацией. Десульфофикация, возбудителем которой являются анаэробные бактерии родов Desulfovibrio и Desulfotomaculum, приводит к восстановле-нию сульфатов до сероводорода, ядовитого для растений: поставщиком водорода служат углеводы, органические кислоты, метан, причем во всех случаях процесс восстановления проходит через следующие промежуточ-ные этапы: 2SO4 ■* H 2SO3 ■* H 2SO2^H 2SO ■* H 2S. Автотрофные бактерии осуществляют в почве процесс окисления недоокисленных минеральных соединений, образующихся в тезультате деятельности гетеротрофов. Основными типами процессов являются нит-рификация, сульфофикация, окисление закисного железа и водорода. Нитрификация. Образующийся в процессе аммонификации аммиак подвергается в почве дальнейшему окислению и переходит сначала в азо-тистую, а затем в азотную кислоту. Окисление аммиачных солей до нитра-тов называется нитрификацией. Процесс нитрификации осуществляется в две фазы: Аммиак окисляется до азотистой кислоты: 2NH3 + 3O2 ■* 2HNO2 + 2H2 O +273 кДж. 161 окислении аммиака до азотистой кислоты принимают участие бак-терии рода Nitrosomonas и других родов. Азотистая кислота окисляется до азотной: 2HNO2 + O2 ■* 2HNO3 + 70 кДж. Окисляют азотистую кислоту до азотной бактерии родов Nitrobacter, Nitrospira, Nitrococcus. Нитрифицирующие бактерии - аэробы и требуют для развития по-стоянного притока кислорода. Они более активны в почвах, хорошо аэри-руемых, близких к нейтральным, содержащих большое количество орга-нических соединений. Поэтому нитрификация протекает интенсивно в дерново-подзолистых почвах, достаточно унавоженных и хорошо обраба-тываемых, а также в черноземных и каштановых почвах, имеющих благо-приятный водный режим. процессе нитрификации образующаяся азотная кислота, соединя-ясь с основаниями, находящимися в почве, дает нитраты, которые исполь-зуются растениями в качестве азотной пищи. При активной нитрификации почве может накопиться в течение года до 300 кг нитратов на 1 га. Нитраты почвы хорошо потребляются растениями, но будучи легко- растворимыми в воде, могут легко вымываться в более глубокие слои поч-вы или за ее пределы. Сульфофикация - это окисление сероводорода, элементарной серы тиосоединений до серной кислоты. Она осуществляется серобактериями родов Beggiatoa, Thiothrix, Thioploga и тионовыми бактериями рода Thio-bacillus. Химическая сущность сульфофикации может быть выраженауравнениями 2H 2S + O2 ■* 2H 2O + 2S + 168 кДж, 2S + 3O2 + 2H 2O ■* 2H 2SO4 + 484 кДж. Выделяющаяся энергия используется серобактериями для усвоения углерода из CO2и синтеза органических веществ. Серобактерии широко распространены в почве, водоемах и имеют большое практическое значение. Серная кислота, получающаяся в резуль-тате деятельности серобактерий, соединяясь с находящимися в почве осно-ваниями, образует сульфаты, которые потребляются зелеными растениями. Окисление железа. Превращение солей закисного железа в окисные происходит при участии железобактерий, широко распространенных в во- 162 доемах и заболоченных почвах. Реакция окисления протекает по уравне-нию 4FeCO3 + 6H 2O +O2 ■* 4Fe (OH)3 + 4CO2 + 160 кДж. Выделяющаяся энергия используется железобактериями для ассими-ляции углерода из CO2 и синтеза органических веществ. Среди этих бакте-рий более широко представлены нитевидные формы (Gallionella, Leptothrix). Азотфиксация. Почвенные бактерии, способные фиксировать азот атмосферы, делятся на 2 группы: свободноживущие в почве аэробные и анаэробные бактерии; клубеньковые бактерии, живущие в симбиозе с растениями. Важнейшим аэробным почвенным организмом, ассимилирующим азот атмосферы, является Azotobacter. К аэробным азотфиксаторам отно-сятся также другие представители семейства Azotobacteriacea, бактерии родов Klebsiella, Erwinia, Pseudomonas. К анаэробным бактериям, которые могут фиксировать атмосферный азот, относятся Clostridium pasteurianum и другие представители рода Clostridium. Клубеньковые бактерии Rhizobium были впервые обнаружены в клу-беньках на корнях бобовых растений М.С. Ворониным (1860 г.). Эти бак-терии проникают в корни бобовых растений и образуют на них вздутия (клубеньки). Они получают сахар от бобовых растений, а его снабжают фиксированным азотом атмосферы. 6.4. Основные принципы индикации и диагностики почв основе принципа биологической диагностики почв лежит пред-ставление о том, что почва как среда обитания составляет единую систему населяющими ее популяциями разных организмов. В зависимости от со-четания природных факторов, определяющих почвообразовательный про-цесс, разные почвы различаются по составу своей биоты, направленности биохимических превращений и содержанию тех химических компонентов, которые являются продуктами этих превращений или их агентами. Биологическая диагностика почв - это определение фактического ипрогнозируемого состояния почвы по ее биологическим свойствам. C од-ной стороны, оно заключается в биоиндикации (биотестировании) свойст-венных почве признаков и тенденций в конкретном месте в конкретное 163 время, то есть в распознавании современного состояния почвы по ее био-логическим свойствам; с другой стороны, в прогнозировании динамично развивающихся в ней процессов и тенденций с определением как долевого, так и суммарного эффектов воздействия на почву факторов естественного и антропогенного происхождения. Биоиндикация (биотестирование)-более узкое понятие,чем биоди-агностика, понятие, предполагающее разовое, точечное измерение (оцен-ку) состояния среды с применением того или иного показателя. Мониторинг почв, то есть наблюдение за состоянием почв во вре-мени, разделяют на диагностическую и прогностическую составляющие. Мониторинг применяется при интегральной оценке состояния почв, эколо-гическом нормировании, уточнении классификации почв, экологической экспертизе и ОВОС, экологическом контроле. Диагностический мониторинг отвечает за опознание почв по состоя-нию их биоты в естественных природных условиях и характеризует их фактическое состояние в настоящий момент времени. Прогностический мониторинг дает представление о перспективе раз-вития намечающихся в почве процессов и выполняется преимущественно в рамках планируемого эксперимента. Для целей биологической диагностики и мониторинга состояния почв и почвенных процессов возможно применение комплекса биологиче-ских показателей, основанных на исследовании почвенной микрофауны (почвенных простейших), альгофлоры, определении биохимической ак-тивности почв. 6.5. Биоиндикация и диагностика почв Ботанические методы фитоиндикации и диагностики почв наи-более хорошо разработаны. Они входят в особый раздел геоботаники - инди-кационную геоботанику, сформировавшуюся в 30-х гг. прошлого столетия. При составлении описания почв используют, как правило, и характе-ристику растительности. Поэтому к настоящему времени накопился уже большой материал по взаимосвязи почв и растений. Методы фитоиндикации применяют при бонитировке почв, почвен-ных свойств и процессов. Например, путем анализа состава и структуры растительных сообществ, распространения растений-индикаторов или оп- 164 ределения индикаторных признаков у отдельных видов растений можно установить: тип почвы, степень ее гидроморфизма, развитие процессов за-болачивания, соленакопления и т. д. Среди растений обнаружены индика-торы на механический и химический состав почв, степень обогащенности питательными элементами, на кислотность или щелочность, глубину про-таивания мерзлотных почв или уровень грунтовых вод. Почвенно-зоологические исследования диагностического профиля стали развиваться позже ботанических. Только в 1950-х гг. почвенная зоо-логия оформилась как самостоятельная область науки. Накопленный к то-му времени фактический материал по распространению в почвах беспо-звоночных, их фаунистическому составу и адаптационным характеристи-кам позволил сформулировать основные принципы зоологического метода диагностики почв. Теоретической предпосылкой применения почвенно-зоологического метода для целей диагностики почв является представле-ние об «экологическом стандарте» вида (М.С. Гиляров), о потребностях каждого вида в определенном комплексе условий среды. Виды с широкой экологической амплитудой (эврибионты) мало при-годны для индикационных целей, тогда как экологически узковалентные виды (стенобионты) служат хорошими индикаторами определенных усло-вий среды и свойств субстрата. Это положение представляет собой общий теоретический принцип в биологической диагностике. Однако использова-ние одного вида для индикации дает мало уверенности в правильности вы-водов. Лучшее условие - исследование всего комплекса организмов, из ко-торых одни могут быть индикаторами на влажность, другие - на темпера-туру, третьи - на химический или механический состав. Чем больше об-щих видов почвенных животных встречается на сравниваемых участках, тем с большей долей вероятности можно судить о сходстве их режимов, а следовательно, и о близости исследуемых почвенных разностей, о единст-ве почвообразовательного процесса. Не все группы почвенных животных равнозначны с точки зрения их диагностической ценности. Менее других полезны микроскопические формы - простейшие и микроартроподы (клещи, ногохвостки). Их пред-ставители отличаются космополитизмом в силу того, что почва для них не выступает как единая среда обитания, а живут они в системе пор, капилля-ров, полостей, которые можно найти в любой почве. Среди простейших до последнего времени только раковинные амебы были использованы в каче- 165 стве показателей почвенных условий, особенно в некоторых гидроморф-ных почвах. Их легче, чем других простейших, определять по строению их раковинок, которые, кроме того, хорошо сохраняются в почве. Наиболь-шая численность тестацид обнаруживается в кислых лесных почвах под хвойными лесами (десятки тысяч в 1 г при биомассе около 10 кг/га) и наи-меньшая - в солонцах. Из микроартропод наиболее хорошо изучены индикаторные свойства панцирных клещей. Все зональные типы почв России четко различаются по численности, биомассе, продуктивности, характеру вертикального рас-пределения, спектрам жизненных форм и по фаунистическому составу на-селения орибатид. Наибольшей величины их численность достигает в поч-вах таежной зоны, а наибольшей продуктивности - в почвах влажных суб-тропиков. Большинство представителей почвенных микроартропод харак-теризуются высокой степенью космополитизма. Состав их комплексов за-висит не только от почвенных условий, но и от характера и флористиче-ского состава растительности. Поэтому перспективно использование ком-плексов микроартропод не столько для диагностики почв, сколько для ин-дикации повреждающих воздействий на почву. Особенно ценны и удобны для индикационных работ в почвоведении комплексы крупных беспозвоночных (дождевые черви, многоножки, ли-чинки насекомых), которые в меньшей степени космополиты, чем про-стейшие и микроартроподы. Ареалы видов крупных беспозвоночных более надежно изучены и характеризуются определенным комплексом почвенно-климатических условий. Есть много примеров индикаторного значения почвенных беспозвоночных. Так, стафилиниды рода В1еdius и чернотелки рода Ве1 ориs показательны для солончаково-солонцового комплекса почв, многоножки-кивсяки, некоторые мокрицы и легочные моллюски служат индикаторами на содержание в почве извести. Это организмы-калькофилы. Вид дождевых червей Осtо1аsium 1асtеиm и некоторые виды проволочни-ков также являются показателями высокого содержания кальция в грунто-вых водах. Для целей установления типа почвы и направления почвообра-зовательного процесса используется не только принцип сравнительного изучения комплекса почвенных животных, но и такие показатели, как профильное распределение беспозвоночных по отдельным горизонтам, их участие в переработке опада. Например, в формировании бурых лесных почв большую роль играют диплоподы, мокрицы, моллюски, энхитреиды, сосредоточенные в лесной подстилке и перерабатывающие опад in situ. 166 серых лесных почвах значительное влияние на формирование профиля оказывают крупные сапрофаги - дождевые черви, связанные с минеральными слоями почвы, куда они вовлекают растительные остатки. Соотношение подстилочных и собственно почвенных крупных форм са-профагов может помочь в некоторых случаях решению вопроса о направ-ленности почвообразовательного процесса. |