Лекции по биологии (1 семестр). Лекция 1 Типы клеточной организации
 Скачать 1.41 Mb. Скачать 1.41 Mb.
|

.
Для всех способов бесполого размножения характерна одна черта – источником наследственной информации служит одна клетка или группа клеток одного родителя. Это ведёт к образованию копии материнского организма. Дочерние особи генетически однообразны, их фенотипы почти одинаковы; поэтому в такой группе особей естественный отбор мало эффективен. Преимущество бесполого размножения состоит в простоте и эффективности: не нужно находить партнера; потомство может оставить любая особь и в любом месте; удачные мутации не теряются, а воспроизводятся в каждом поколении. При переходе к половому размножению эти преимущества утрачиваются. Однако в результате оплодотворения происходит комбинирование признаков мужских и женских организмов, чем обеспечивается генетическое разнообразие потомства. Возникает новый неисчерпаемый источник наследственной изменчивости, который делает естественный отбор эффективным. Появление полового размножения – важное эволюционное событие. Оно придаёт популяции пластичность, что особенно необходимо в изменчивой среде.
Разделение половых клеток на яйцеклетки сперматозоиды, половых желез на яичники и семенники, а особей на самок и самцов – называется половым диморфизмом. Генетический аспект: в основе полового диморфизма лежат различия в хромосомном наборе мужских и женских и особей. Морфофизиологичекий аспект: в связи с различием в хромосомном наборе мужских и женских и особей организмы имеют свое морфологическое строение, влияющее на физиологические функции и процессы. Эндокринный аспект: мужские (тестостерон) и женские (эстрогены) половые гормоны оказывают различное воздействие на человека, что приводит к появлению вторичных половых признаков. Поведенческий аспект: наличие мужских и женских вторичных половых признаков и интенсивная выработка половых гормонов в период полового созревания обуславливает различное поведение самцов и самок.
Партеногенез – это разновидность полового размножения, когда развитие организма происходит из неоплодотворенной яйцеклетки. Партеногенез подразделяется естественный и искусственный. В свою очередь естественный партеногенез делится на облигатный (обязательный) и факультативный (необязательный). При облигатном партеногенезе яйца способны только к партеногенетическому развитию. Такой партеногенез наблюдается у некоторых популяций скальных ящериц. При факультативном партеногенезе яйца могут развиваться и посредством партеногенеза и в результате оплодотворения. Такой партеногенез наблюдается у пчел, когда трутни развиваются из неоплодотворенных яиц, а матка и рабочие пчёлы – из оплодотворённых яиц. Искусственный партеногенез вызывают действием на яйца различных раздражителей: уколом иглой, обработкой растворами сильных кислот, резким охлаждением и особенно нагревом. С помощью искусственного партеногенеза обычно удается получать лишь начальные стадии развития организма; полный партеногенез достигается редко, хотя известны случаи полного партеногенеза даже у позвоночных животных (лягушка, кролик). Способ массового получения полного партеногенеза, разработанный (1936) для тутового шелкопряда Б.Л. Астауровым, основан на точно дозированном кратковременном прогреве (до 46ºС в течение 18 мин) извлеченных из самки неоплодотворенных яиц. Этот способ дает возможность получать у тутового шелкопряда особи только женского пола, наследственно идентичные с исходной самкой и между собой. Таких особей можно потом размножать посредством партеногенеза неограниченно долго.
Человек – раздельнополое млекопитающее. Имеет разделение половых клеток на яйцеклетки и сперматозоиды, половых желез на яичники и семенники, а также различия в хромосомном наборе ♀ и ♂ особей. Характерна медленная смена поколений, низкая плодовитость (обычно 1–2 ребенка, не сравнить с дрозофилой, или тараканом). ЛЕКЦИЯ 4 Половые клетки. Гаметогенез. Мейоз.
Яйцеклетка – это крупная, округлая, неподвижная клетка. Размеры варьируют, диаметр у млекопитающих 100 – 200 мкм (микрометров). Это типичная эукариотическая клетка. В ней выделяют: оболочку, цитоплазму и гаплоидное ядро. В цитоплазме находятся органоиды и включения в виде желтка. В желтке много белка, а также находится жироподобное вещество лецитин. В зависимости от количества и распределения желтка по цитоплазме выделяют следующие типы яйцеклетки. 1. Изолецитальные – желтка мало и он равномерно распределен по цитоплазма. Ядро в них располагается ближе к центру (черви, моллюски, ланцетник, млекопитающие). 2. Телолецитальные – желтка много и он находится на вегетативном (растущем) полюсе клетки, а ядро будет на противоположном анимальном полюсе (рыбы, амфибии, птицы) а) умеренно телолецитальные яйцеклетки содержат среднее количество желтка (осетровые рыбы, земноводные). б) резко телолецитальные яйцеклетки содержат очень много желтка, занимающего почти весь объем клетки (некоторых рыбы, пресмыкающиеся, птицы, яйцекладущие млекопитающие). 3. Центролецитальные – желтка много и он находится в центре клетки (насекомые). Все яйцеклетки имеют плазматическую мембрану. Яйцеклетки многих животных кроме плазматической мембраны имеют дополнительные защитные оболочки. Первичная оболочка, или желточная. Она образуется в результате жизнедеятельности самой яйцеклетки. У млекопитающих её называют блестящей оболочкой или зона пеллюцида. Zona pellucida Вторичная оболочка (хорион у млекопитающих) образуется в результате жизнедеятельности фолликулярных клеток, которые окружают яйцеклетку в яичнике. Третичная оболочка образуется при прохождении яйцеклетка по яйцеводу. В яйцеклетке птиц это белковая оболочка, подскорлуповая, и скорлуповая оболочки
Сперматозоид человека (50мкм), состоит из головки, шейки, средней части и хвостика. Головка содержит ядро с ДНК, передняя часть головки содержит акросому – структуру, выделяющую ферменты для растворения оболочки яйцеклетки. Это видоизменённый комплекс Гольджи, содержащий лизосому. В шейке содержатся две центриоли: одна участвует в образовании веретена деления, а другая – оси хвоста. Средняя часть содержит митохондрии, обеспечивающие сперматозоид энергией во время движения. Хвостик – органоид движения.
Гаметогенез – процесс образования половых клеток. Он делится на сперматогенез – развитие мужских половых клеток – и овогенез – развитие женских половых клеток. В гаметогенезе выделяют 5 периодов: обособление, размножение, рост, созревание и формирование. 1. Обособление – первичные половые клетки обособляются от соматических клеток. Однако, они содержат диплоидный набор хромосом, их генетическая формула 2n2c. 2. Размножение – первичные половые клетки (сперматогонии или овогонии) делятся митозом. Задача этого периода увеличить число первичных половых клеток. Генетическая формула 2n2c. 3. Рост – сперматогонии и овогонии накапливают питательные вещества и увеличиваются в размерах. Теперь они называются сперматоциты I порядка и овоциты I порядка. В конце периода происходит репликация хромосом (2п2с 2n4с). 4. Созревание (мейоз) – происходит два последующих деления, между которыми нет интерфазы, и, следовательно, нет удвоения ДНК. Набор хромосом в клетках уменьшается в два раза, а набор хроматид уменьшается в 4 раза (мейоз I: 2n4с1n2с, мейоз II: 2n2с1n1с). 5. Формирование – клетки приобретают специфическое строение, обеспечивающее выполнение их функции. Этот период характерен только для сперматогенеза. Отличия овогенеза от сперматогенеза СПЕРМАТОГЕНЕЗ ОВОГЕНЕЗ
Мейоз – это два последовательных деления клетки, приводящие к уменьшению числа хромосом вдвое. Каждое деление мейоза включает в себя 4 фазы: профаза, метафаза, анафаза и телофаза. Перед мейозом в конце периода роста происходит репликация ДНК, клетка имеет набор хромосом 2n4с. Профаза I . Это самая продолжительная фаза мейоза I, во время которой происходят события, отличающие мейоз от митоза. Профаза подразделяется на пять стадий, в неё вступают гаметоциты 1 порядка (2n4с)
Механизм конъюгации:
Кроме того, в профазу 1 центриоли расходятся к разным полюсам клетки – образуется веретено деления. Разрушаются ядрышки, ядерная оболочка. В области центромеры с одной стороны каждой хромосомы образуются кинетохоры, от них отходят кинетохорные нити. Метафаза 1. Конъюгированные хромосомы (биваленты) располагаются по экватору клетки, образуя метафазную пластинку. Заканчивается формирование веретена деления. В отличие от митоза нить веретена от каждой хромосомы направлена только к одному из полюсов. Это происходит потому, что из-за конъюгации каждая хромосома имеет только один кинетохор. Анафаза 1. Гомологичные хромосомы, состоящие каждая из 2-х хроматид, отходят к противоположным полюсам. Расходятся хромосомы, а не хроматиды, т.к. центромера не делится, а хромосомная нить веретена тянется лишь к одному полюсу. Очень важно, что расхождение гомологичных хромосом происходит случайным образом. Поэтому к каждому полюсу отходит случайное число отцовских или материнских хромосом. На каждом полюсе находится в 2 раза меньше хромосом, чем было их в клетке до начала деления. Причем эти хромосомы качественно другие: большая часть каждой хромосомы – исходная хромосома; меньшая часть представлена заменённым участком гомологичной хромосомы. Так как гомологичные хромосомы – это отцовская и материнская хромосомы, можно сказать, что в результате кроссинговера образуются комбинированные хромосомы. В них содержатся новые комбинации отцовских и материнских генов, которые будут служить материалом для эволюционного процесса. Tелофаза I – образуются 2 клетки (сперматоциты II-го порядка, овоциты II-го порядка) с набором 1n2с. Хромосомы деспирализуются, образуется ядерная оболочка, исчезает веретено деления, делится цитоплазма (цитокинез). Так как набор хромосом уменьшается вдвое, первое мейотическое деление называется редукционным. После короткой интерфазы, во время которой не происходит репликации ДНК (т.к. отсутствует S-фаза), наступает мейоз II. Он называется эквационным, или уравнительным. Мейоз II напоминает митоз, но особенность в том, что набор хромосом клеток, вступающий в мейоз II гаплоидный, и расходящиеся хроматиды содержат новую комбинацию генов по сравнению с хроматидами исходной клетки. Итак, при сперматогенезе и овогенезе из одной исходной диплоидной клетки (2n4с) образуются четыре клетки с гаплоидным набором хромосом (1n1с), причем эти хромосомы содержат новую комбинацию генов. Общая характеристика половых клеток, или гамет.
7. Биологическое значение мейоза. 1. мейоз обеспечивает постоянный для каждого вида организмов набор хромосом и постоянное количество ДНК. Если бы в процессе мейоза не происходило уменьшение числа хромосом, то в каждом следующем поколении после оплодотворения число хромосом возрастало бы в 2 раза. Благодаря мейозу, зрелые гаметы получают гаплоидное число хромосом, а при оплодотворении восстанавливается свойственное данному виду диплоидное число хромосом; 2. мейоз обеспечивает генетическое разнообразие гамет. Это достигается, благодаря двум явлениям: кроссинговеру и независимому расхождению мужских и женских хромосом в мейозе - I и хроматид в мейозе - II. Эти явления лежат в основе комбинативной изменчивости, поставляющей материал для естественного отбора. ЛЕКЦИЯ 5 Законы Г. Менделя. 1. Наследственность и наследование, их сущность. Генетика – наука о закономерностях наследственности и изменчивости. Наследственность – способность родительских форм передавать при размножении свои признаки потомству. Наследственность консервативна, она сохраняет уже возникшие черты и свойства организмов на протяжении многих поколений. Материальной основой наследственности является наличие генов в хромосомах и закономерности поведения их в процессе гаметогенеза и размножения. Наследование – это способ передачи наследственных признаков в ряду поколений. Наследование является внешним проявлением наследственности и именно с явлением наследования тех или иных признаков имеет дело врач. 2. Ген как единица функционирования наследственного материала. Современная генетика рассматривает ген как единицу функционирования наследственного материала. Это означает, что передача генов в ряду поколений обеспечивает наследование потомками признаков родителей. Ген – это участок молекулы ДНК, содержащий последовательность нуклеотидов, которая кодирует последовательность аминокислот в одиночной полипептидной цепи, либо кодирует последовательность нуклеотидов в тРНК или рРНК. 3. Свойства генов: стабильность, аллельное состояние, специфичность, дискретность. Стабильность. Гены в ряду поколений не изменяются. Аллельное состояние. Аллельные гены – это гены, которые находятся в одинаковых локусах гомологичных хромосом. Они отвечают за развитие альтернативных признаков (за разное выражение данного признака). Специфичность – один ген отвечает за развитие одного признака. Дискретность – за развитие разных признаков отвечают разные гены, находящиеся в разных хромосомах. Примечание. Эти свойства генов были описаны в самом начале 20в в рамках представлений классической генетики. 4. Понятие о гомозиготности и гетерозиготности. Организмы с одинаковыми аллелями одного гена называются гомозиготными, или гомозиготами. Гомозигота может быть доминантной (АА) или рецессивной (аа). Организмы, имеющие разные аллели одного гена: один доминантный, другой рецессивный, называются гетерозиготными, или гетерозиготами (Аа). В результате мейоза гомологичные хромосомы, а с ними и аллельные гены расходятся в разные гаметы. Так как у гомозиготной особи оба аллеля одинаковы, она образует один тип гамет. Гетерозиготная особь образует 2 типа гамет – один тип с доминантным аллелем, другой – с рецессивным аллелем 5. Гибридологический анализ – основной метод генетики. Основной метод, используемый Г. Менделем, – гибридологический. Гибрид – особь, полученная в результате полового размножения. Так как потомок сочетает признаки обоих родителей, то по наличию у него определённых признаков можно судить о наличии у его родителей соответствующих генов. Г. Мендель использовал гибридологический метод в отличие от своих предшественников при соблюдении следующих условий: 1. в каждом поколении вёлся учёт по каждой паре альтернативных (взаимоисключающих) признаков без учёта других различий скрещиваемых организмов. Таким образом, Г. Мендель решал задачу с одним неизвестным, а его предшественники решали задачу со многими неизвестными, т.к. учитывали наследование всей совокупности признаков организма; 2. проводился строгий количественный учёт гибридов, различающихся по отдельным парам альтернативных признаков, в ряду последовательных поколений; 3. проводился индивидуальный анализ потомства от каждого гибридного организма. 6. Открытие Г. Менделем законов независимого наследования. Моногибридное скрещивание. Единообразие гибридов первого поколения. Единообразие гибридов первого поколения было установлено при моногибридном скрещивании гороха, т.е. скрещивания, при котором изучалось наследование одного признака – цвета горошин. Горошины могли иметь либо жёлтый, либо зелёный цвет (это альтернативные признаки). Горох – самоопыляемое растение, причем опыление происходит в бутоне. Это устраивало Г. Менделя, т.к. позволяло ему целенаправленно проводить скрещивание растений путём искусственного опыления. Он скрестил гомозиготные растения, имеющие зелёные и жёлтые семена. Независимо от того, какой цвет семян имело материнское растение, гибридные семена были жёлтыми. Таким образом, у гибридов первого поколения проявился признак только одного родителя. Это доминантный признак – жёлтый цвет семян; рецессивный признак – зелёный цвет семян – как бы исчезал. Р  Г  F1 Аа, Аа, Аа, Аа Итак, все потомки имеют одинаковый генотип, а т.к. фенотипически проявляется только доминантный аллель – все потомки имели семена только жёлтого цвета. Гибриды первого поколения единообразны по генотипу, а, следовательно, и по фенотипу. Можно так сформулировать правило единообразия: "При скрещивании гомозиготных особей, отличающихся друг от друга по одной паре альтернативных признаков, все гибриды первого поколения будут иметь признак одного из родителей, и поколение по данному признаку будет единообразным".    7. Закон расщепления. Доминантность и рецессивность. На следующий год Г. Мендель скрестил (самоопылением) гибриды первого поколения между собой. Осенью при подсчёте семян оказалось, что из 8023 семян 6022 были жёлтыми, 2001– зелёными, т.е. соотношение 3:1. Итак, во втором поколении проявляется признак зелёной окраски, и он присущ 1/4 части потомства. Такое явление Г. Мендель назвал расщеплением признаков и сформулировал закон расщепления: "В потомстве, полученном от скрещивания гибридов первого поколения, анализируемых по одной альтернативной паре признаков, наблюдается явление расщепления: 3/4 части особей второго поколения несёт доминантный признак, 1/4 часть – рецессивный". Р Г  F2 АА, Аа, Аа, аа При скрещивании между собой гибридов второго поколения Г. Мендель обнаружил в их потомстве следующее: в потомстве зелёных семян (аа) расщепления не наблюдалось; 1/3 растений, выросших из жёлтых семян (АА), произвела только жёлтые семена; 2/3 растений, выросших из жёлтых семян (Аа), произвела жёлтые и зелёные семена в соотношении 3:1. Таким образом, Г. Мендель впервые установил факт, свидетельствующий о том, что растения, сходные по внешнему виду, могут резко отличаться по наследуемым свойствам. Гомозиготы (АА) не давали расщепления, а гетерозиготы (Аа) давали расщепление по фенотипу в следующих поколениях в отношении: три части особей с доминантным признаком к одной части особей с рецессивным признаком. Расщепление по генотипу сложнее: I часть доминантных гомозигот (АА); 2 части гетерозигот (Аа); I часть рецессивных гомозигот (аа). 8. Закон чистоты гамет. Анализирующее скрещивание. Для объяснения закона расщепления Г. Мендель выдвинул гипотезу «чистоты гамет». Её суть в следующем. Любой организм содержит в каждой соматической клетке два аллеля любого гена, расположенных в гомологичных хромосомах. В гаметах же содержится по одному аллельному гену. Это происходит потому, что гомологичные хромосомы при мейозе расходятся к разным полюсам делящейся клетки и попадают в разные половые клетки. Следовательно, гаметы имеют из пары гомологичных хромосом только одну, и, соответственно, только один аллельный ген. Таким образом, гаметы «чисты» от другого аллельного гена. Гибрид, полученный от слияния гамет, содержит оба аллельных гена, но фенотипически всегда проявляется лишь доминантный аллельный ген, а рецессивный ген проявляется только в гомозиготном состоянии. Отсюда становится понятным, что из поколения в поколение гены передаются не меняясь. Иначе как объяснить, то во втором поколении после скрещивания растений с жёлтыми горошинами снова появились растения с зелёными горошинами. Для доказательства гипотезы «чистоты гамет» Г. Мендель провёл анализирующее скрещивание, т.е. скрещивание гетерозиготной особи и гомозиготной рецессивной особи. Г. Мендель рассуждал так: если гетерозиготная особь образует гаметы, в которых содержатся оба аллельных гена, всё потомство будет жёлтым: Р Г   F1 Аааа Если же аллельные гены попадают в разные гаметы, то в потомстве должно быть расщепление в соотношении: 50% особей с доминантным признаком и 50% с рецессивным признаком: Р Г  F1 Аа, аа Эксперимент подтвердил справедливость второго варианта: гетерозигота даёт два типа гамет. Следовательно, гипотеза «чистоты гамет» верна. С открытием мейоза «гипотеза чистоты гамет» получила цитологическое подтверждение. В настоящее время, анализирующее скрещивание используется для установления гомозиготности или гетерозиготности организма по доминантному признаку. Известно, что рецессивный признак проявляется фенотипически только при гомозиготности рецессивного гена (аа), а доминантный признак проявляется как при гомозиготности доминантного гена (АА), так и при гетерозиготности (Аа). Анализирующее скрещивание заключается в том, что особь, генотип которой необходимо выяснить, скрещивается с особью, гомозиготной по рецессивному признаку. Полученные гибриды анализируются. 1 вариант 2 вариант Р Г   Г Г F1 Аа F1 Аа, аа Как видим из схемы, при анализирующем скрещивании в потомстве гомозиготной доминантной особи нет расщепления, гетерозиготная особь даёт расщепление в соотношении 1:1. Иначе говоря, наличие в потомстве первого поколения расщепления говорит о гетерозиготности организма по исследуемому признаку. 9. Дигибридное и полигибридное скрещивание. Закон независимого наследования признаков. Дигибридное скрещивание – это скрещивание, при котором изучается наследование двух пар признаков, причем гены, контролирующие эти признаки, лежат в разных хромосомах.
Г. Мендель брал растения с семенами жёлтого цвета и гладкой формы и скрещивал их с растениями, дающими семена зелёного цвета и морщинистой формы. При скрещивании гомозиготных особей получилось единообразное по фенотипу потомство – все семена были жёлтые и гладкие. Р 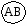 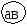 Г F 1 АаВв Затем Г. Мендель скрестил гибриды первого поколения между собой (самоопыление). В их потомстве наблюдалось расщепление признаков: 9 частей семян жёлтых гладких, 3 части семян жёлтых морщинистых, 3 части семян – зелёных гладких и I часть семян – зелёных морщинистых. Р 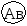 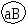 Г 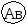 Для записи дигибридного скрещивания удобно пользоваться решеткой Пеннета:
Расщепление по фен-пу: 9 частей семян Ж.Г.: 3 части семян Ж.м.: 3 части семян з.Г. : I часть семян з.м. Затем Г. Мендель проанализировал расщепление отдельно по цвету и форме семян. Оказалось, что по окраске на 3 части жёлтых семян пришлась I часть зелёных. По форме наблюдалось такое же расщепление: 3 части семян гладких на I часть морщинистых. Г. Мендель делает вывод: дигибридное скрещивание есть 2 моногибридных скрещивания, идущих независимо друг от друга. Математически это можно выразить так: (3+I)2 = 9+3+3+1. На основе этого вывода Г. Мендель формулирует закон независимого наследования: "Расщепление по каждой паре признаков идет независимо от других пар признаков". 10. Условия менделирования признаков. Менделирующие признаки человека. У человека много признаков, которые при наследовании подчиняются законам Менделя. Такие признаки называются менделирующими признаками. Это могут быть как нормальные, так и патологические признаки. Условия менделирования признаков.
ЛЕКЦИЯ 6 Сцепленное наследование признаков. Наследование признаков, |

