лекции по генетике. Лекция Законы наследования
 Скачать 211.76 Kb. Скачать 211.76 Kb.
|

|
Хромосомная теория наследования. Была сформулирована на основе законов Менделя в основном благодаря группе ученых под руководством Томаса Ханта Моргана. К концу 19 века стали известны закономерности митоза и мейоза. Молодой американский ученый Сеттен предположил, что менделевские наследственные факторы локализованы в хромосомах, однако первым это смог доказать Морган. В современном виде можно выделить следующие положения хромосомной теории наследственности:
Сцепленное с полом наследование. Первые доказательства хромосомной теории наледования было получено Морганом на плодовой мушке дрозофиле. Этот объект является наиболее удобным в генетике, т.к. существует много вариантов признаков и поэтому можно легко получить гомозиготные линии, которые будут отличаться по ряду альтернативных признаков. Мушка образует большое количество потомков срок между двумя генерациями не велик, всего 10 дней. Заслуга Моргана состоит в том, что он первым ввел этот объект в генетику .Еще до Моргана было известно, что представители разных полов отличаются друг от друга по набору половых хромосом и возможны два варианта наборов:
У дрозофилы гомогаметный пол женский, гетерогаметный – мужской. Морган вывел гомозиготные линии мух с красными глазами и с белыми глазами и проводил гибридизацию в том порядке, в котором это делал Мендель. Он предположил, основываясь на работах Сеттена, что ген, определяющий цвет глаз, локализован в X-хромосоме, а в Y-хромосоме его нет. Благодаря своим работам Морган смог это доказать.
Подтверждает первый закон Менделя, т.е. закон единообразия первого поколения. Р: ♀ХАХа +♂ХАY F: ♀ХАХА: ХАХа ♂ХаY: ХАY Подтверждается закон расщепления. Однако Морган обратил внимание на то, что белоглазыми в данном скрещивании были только самцы, следовательно, Морган делает вывод, что ген, определяющий цвет глаз, локализован в Х-хромосоме, а в Y-хромосоме его нет. Морган проводит рецепрокное скрещивание. Р: :♀ХаХа+♂ХАY F:♀ХАХа ♂ХаY 1:1 Нарушается закон единообразия первого поколения. Наблюдается крисс-кросс наследование, т.е. дочери наследуют признак отцов, а сыновья – матерей. Явление крисс-кросс наследования это нарушение первого законы Менделя, которое возможно благодаря тому, что признак локализован в Х-хромосоме. Р: ♀ХАХа +♂ХаY F: ♀ХаХа: ХАХа ♂ХаY:ХАY 1:1 Полученные результаты Морган объяснял тем, что ген локализован в Х-хромосоме, а в Y-хромосоме его нет. Т.о. Морган подтверждает, что гены находятся в половых хромосомах. Сцепленное с полом наследование – это наследование генов, расположенных в половых хромосомах. Существует три варианта наследования сцепленного с полом:
1). У человека приблизительно 200 генов локализовано в Х-хромосоме, которых нет в Y-хромосоме. Некоторые из них имеют медицинское значение:
Для млекопитающих характерен ряд особенностей при полностью сцепленном с полом наследовании. Одна из Х-хромосом у женского гомогаметного пола в клетке сильно спирализуется, превращается в тельце Барра и инактивируется, т.е. в соматических клетках у самок млекопитающих функционирует только одна Х-хромосома. Процесс инактивации случаен, поэтому полностью сцепленное наследование у млекопитающих имеет свои особенности. Классическим примером является наследование окраски шерсти у кошек.
Иногда встречаются черепаховые самцы, которые имеют генотип ХХY, они стерильны. У человека также возможна инактивация одной из Х-хромосом, что приводит к определенному мозаицизму признаков. Например, существует болезнь, связанная с отсутствием потовых желез, у мужчин потовые железы могут полностью отсутствовать, а у женщин могут располагаться мозаично на теле. Мозаичность может проявлятся у женщин при дальтонизме. Женщина может быть дальтоником на один глаз, либо дальтонизм проявляется на уровне сетчатки. Полностью сцепленное с полом наследование у бабочек и птиц имеет свои особенности, т.к. у них гомогаметный пол – мужской, а гетерогаметный – женский.
У некоторых насекомых сцепленное с полом наследование имеет свои особенности, связанные с тем, что представители разных полов различаются по количеству хромосом. Самки таких насекомых имеют набор ХХ, а самцы ХО
F: ♀ХХ: ♂ХО 1:1 2). Частично сцепленное с полом наследование. В данном случае признаки наследуются от гетерогаметного пола к гетерогаметному, а от гомогаметного к гомогаметному, т.е. крисс-кросс наследования не наблюдается. Биологический смысл частично сцепленного с полом наследования заключается в том, что это один из механизмов полового диморфизма. Ген, который контролирует толщину эмали на зубах располагается и в Х и в Y хромосомах. из-за того, что одна из Х-хромосом у гомогаметного пола спирализуется, эмаль на зубах толще у мужчин.
3). Голандрическое наследование, при котором аллели локализованы в Y-хромосоме. В Y-хромосоме находится не много генов, но как правило они определяют развитие первичных и вторичных мужских половых признаков. Голандрическое наследование лежит в основе полового деморфизма. В Y-хромосоме локализованы следующее гены: SRY – этот ген отвечает за синтез белкового фактора, который стимулирует развитие семенников SPY – ген, который отвечает за сперматогенез HYA - ген, который отвечает за синтез белка, входящего в состав плазмы крови Наследование идет по следующему принципу: от гетерогаметного пола к гетерогаметному полу.
Существуют признаки, которые могут быть локализованы в аутосомах, но которые в норме проявляются только у одного пола. Это признаки ограниченные полом, например, оволосенение, яйценоскость у кур, этот ген лаколизован в аутосомах. Существуют признаки, зависящии от пола, эти признаки по-разному проявляются у представителей разных полов, например, у человекав ранняя алопеция.
Генетика пола. Пол – это набор определенных признаков, которые обеспечивают размножение. Определение пола у разных животных осуществляется на разных этапах онтогенеза:
на ранних этапах развития зародыш бисексуален. Есть гены для развития как женских, так и мужских половых признаков. Решающее значение в этом случае имеет баланс или соотношение этих генов. У женского пола в Х-хромосоме есть гены, определяющие развитие женских половых признаков. В Y-хромосоме есть гены SRY и SPY. Существуют женщины генотипом ХY, у которых данные гены не работают. До восьми недель беременности у эмбрионов формируется совершенно одинаковая система первично половых признаков. 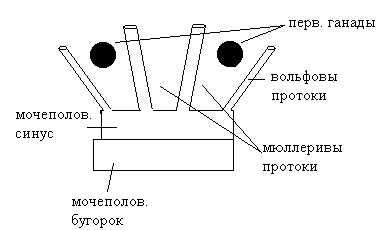 После восьми недель беременности начинаются различия, если плод генотипом XX, то под действием гормонов матери первичные гонады преобразуются в яичники, которые как железа внутренней секреции начинают работать только после периода полового созревания, следовательно, развитие женского эмбриона определяется только гормонами матери. Под их влиянием дегенерируют вольфовы протоки, а мюллеровы превращаются в яйцеводы или в фаллопиевы трубы. Из мочеполового синуса развивается шейка матки, матка и верхняя часть влагалища. Из мочеполового бугорка развивается нижняя часть влагалища и наружные гениталии. Гормоны матери действуют на гипоталамус зародыша и детерминируют поведение по женскому половому типу после полового созревания. У зародышей с генотипом XY из первичных гонад развиваются семенники, которые сразу же начинают функционировать как эндокринные железы, т.е. у эмбриона начинает синтезироваться тестостерон. В результате мюллеровы протоки дегенерируют, из вольфовых развиваются семяпроводы, из мочеполового синуса и мочеполового бугорка - семяизвергательный канал и канал уретры, из мочеполового бугорка – мошонка и половой член. Тестостерон действует на гипоталамус зародыша и после периода полового созревания определяется поведение по мужскому половому признаку. С медицинской точки зрения существуют следующее уровни определения пола:
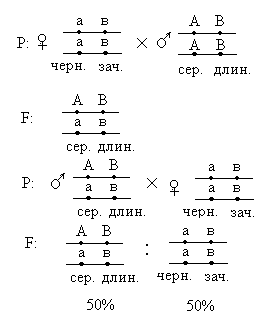
Лекция 2. Сцепленное наследование Сцепленным наследованием называется наследование, при котором признаки локализованы в одной хромосоме и наследуются совместно. Сцепленным наследованием занимался Морган на дрозофиле, для которой было известно, что гаплоидное число хромосом равно 4, а альтернативных пар признаков намного больше, поэтому Морган предположил, что гены, контролирующие некоторые признаки, локализованы в одной хромосоме. Морган исследовал два признака: окраску тела и длину крыльев, и провел два рецепрокных скрещивания.
Морган делает вывод: раз наследуются только родительские комбинации, то данные признаки сцеплены, они локализованы в одной хромосоме и образуют группу сцепления. Это является доказательством как 1ого, так и 2ого положений хромосомной теории. В данном случае наблюдается так называемое абсолютное сцепление (крайне редкий вариант). Морган проводит рецепрокное анализирующее скрещивание.  Так как в результате рецепроконого скрещивания получились рекомбинантные классы, то можно сделать вывод, что сцепление между генами неполное или частичное. В этом случае появление рекомбинантных классов можно объяснить с точки зрения того, что между гомологичными хромосомами в процессе мейоза происходит кроссинговер. 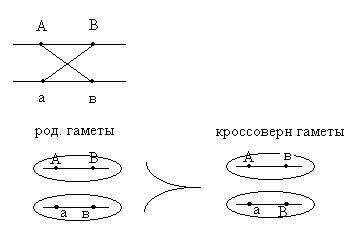 К моменту данных опытов мейоз был известен и было известно, что в профазе мейоза гомологичные хромосомы коньюгируют с образованием бивалентов. Однако кроссинговер был доказан Морганом теоретически и только через 20 лет был обоснован цитологически. Родительских гамет всегда больше чем кроссоверных, причиной этого является то, что кроссинговер при мейозе идет не в 100% случаев, можно учитывать только вариант нечетного кроссинговера, так как результаты четного кроссинговера в потомстве будут незаметны. 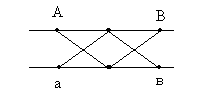 Поэтому чистота кроссинговера теоретически не может превышать 50%. Различие двух реципрокных скрещиваний объясняется тем, что у самок дрозофилы кроссинговер проходит, а у самцов он не идет никогда, поэтому в первом варианте наблюдается редкий вариант абсолютного сцепления. В целом частота кроссинговера у гомогаметного пола выше, чем у гетерогаметного пола. Таким образом, этими опытами Морган доказал, что гены лежат в хромосомах и гены, локализованные в одной хромосоме, образуют группу сцепления и могут наследоваться совместно. Частота кроссинговера и, следовательно, частота появления кроссоверных гамет будет зависеть от расстояния между генами А – В – х% В – С – y % А – С – z % Если анализировать попарное сцепление генов, то можно определить, что гены лежат в хромосоме линейно. 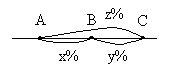 Х%+ Y %= Z % На основании линейного положения хромосом можно строить генетические карты хромосом. На них хромосомы изображены в виде линий, на которых указываются гены и расстояния между генами. Один процент кроссинговера является генетической единицей расстояния на такой карте. Было принято считать, что 100% кроссинговера равняется 1 Моргану. 100% - 1 Морган 1% - 1 Морганида помимо генетических карт хромосом существуют цитологические карты хромосом, причем карты коллинеарны, то есть для них характерен одинаковый порядок генов. Цитологические карты получают при дифференцировочной окраске хромосом. 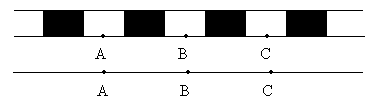 Существуют варианты сцепления трех генов, причем ситуация, когда между всеми генами происходит кроссинговер. Существует общий алгоритм решения такой задачи. 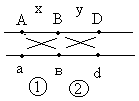 2 n =8 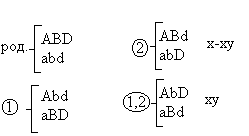 род. 1-(х(1-у)+(у(1-х)=1-х(1-у)-у(1-х)=1-х+ху-у+ху=1-х-у+2ху Лекция 3. Генотип и фенотип Фенотип – это совокупность всех или анализируемых признаков, характерных для данного организма. Фенотип развивается на основе внутренних факторов, материнских факторов и за счет влияния факторов внешней среды. К внутренним факторам относится наследственная информация, которую на уровне вида можно определить как геном, а на уровне организма понятием генотип (совокупность аллелей). Гены в организме представлены в определенном физико-химическом состоянии, которое определяется последовательностью нуклеотидов. Конкретной последовательностью нуклеотидов является аллель. Как правило, для данного вида организмов характерны различные варианты аллелей, то есть фенотип определяет явление множественного аллелизма. Множественный аллелизм заключается в том, что различные аллели контролируют развитие более чем двух вариантов конкретного признака (конкретный вариант признака называется феном). Причиной множественного аллелизма является внутригенная мутация, внутригенный кроссинговер. Классическим примером множественного аллелизма является наследование групп крови у человека. Гены в системе АВО отвечают за синтез различных галактозилтрансфераз. Множественный аллелизм характерен для окраски глаз у дрозофилы. С abr – абрикосовый C cr - коралловый C + - красный C ch - вишневый C eor - эозинофиловый C – белые Множественный аллелизм характерен для окраски шерсти у многих млекопитающих. Существует ген С, которые контролирует структуру фермента тирозолазы, субстратом для этого фермента является тирозин, который под действием фермента превращается в пигмент меланин. Существует два варианта меланина: Эумеланин Феомеланин Существуют различные аллели данного гена: С - черный C ch - шиншилловая C h – гималайская С – белый Множественный аллелизм лежит в основе генетического полиморфизма популяции. Множественный аллелизм существует у растений в основном у перекрестно опыляемых, причем существуют механизмы, которые поддерживают этот генетический полиморфизм. К таким механихмам относится явление самостерильности у покрытосемянных растений, у них существует ген S , который определяет проростание пыльцевой трубки. Существует несколько аллелей генов S ( S 1, S 2, S 3…). 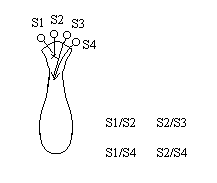 Влияние генотипа на фенотип как правило очень сложное и один ген может влиять не на один, а на целую группу признаков. Такое явление получило название плейотропия или множественное действие генов. Различают два варианта плейотропии:
при первичной плейотропии патологичен продукт гена, но это влияет на множество признаков в организме. Синдром Марфана, болезнь, связанная с патологией в синтезе каллогена. В результате патологично большинство каллогеновых структур, в организме наблюдается деформация скелета, рост, как правило, выше среднего, непропорционально удлинены конечности, арахнодактелия, патологии суставных сумок, различные пороки сердца, патологии органов зрения. Вторичная плейотропия. Классическим примеро является серповидноклеточная анемия. Эта аномалия вызвана одной аминокислотной заменой в В-цепи гемоглобина, в результате гемоглобин плохо связывается с кислородом и ткани накодятся в постоянном дефиците кислорода. Это доминантное заболевание, гомозиготы не выживают, у гетерозигогт наблюдается увеличение селезенки, гипертрофия сердца, гипертрофия красного костного мозга. Гетерозиготы оказываются в некоторых условиях более жизнеспособны, чем гомозиготы по рецессиву (устойчивость к малярии). Патологичные гены не всегда проявляются в фенотипе, причем они могут проявляться с разной степенью выраженности. Вероятность проявления данного аллеля в фенотипе называется пенетрантностью. Если аллель всегда проявляется в фенотипе, то говорят о полной пенетрантности, если аллель не проявляется в фенотипе в различных случаях, то говорят о неполной пенетрантности. Например, полидактилия – это болезнь с неполной пенетрантностью. Из 100 носителей аллелей она проявляется в фенотипе с разной степенью выраженности. Например, полидактилия – это появление пятого пальца на всех конечностях, либо только на одной, в виде нормально развитого пальца, либо только в виде зачатка фаланги, в этом случае говорят о вариабельной экспрессивности развития признаков. Экспрессивность развития признаков бывает константной, если данный аллель всегда проявляется в фенотипе. Влияние генотипа на фенотип проявляется в явлении взаимодействия генов. Лекция 4. Взаимодействие генов Выделяют две крупные ситуации:
причем генотип однозначно определяет фенотип не во всех ситуациях, а только в ситуации промежуточного наследования и кодоминирования. Полное доминирование характерно для генов, которые контролируют структуру ферментов, например, ген IА полностью доменирует над геном I0. промежуточное наследование характерно для структурных белков, белков-рецепторов, белков-переносчиков, для ферментов со средней активностью, классическим примером является наследование структуры волоса. Кодоминирование является наиболее встечающимся вариантом наследования на биохимическом уровне. Классическим примером кодоминирования является проявление 4 группы крови в системе АВО.
 Выделяют три варианта взаимодействия неаллельных генов:
комплиментарность – это такой тип взаимодействия неаллельных генов, при котором наличие в генотипе доминантных аллелей всех взаимодействующих генов определяет развитие самостоятельного признака. Классическим примером является синтез глазных пигментов у дрозофилы. 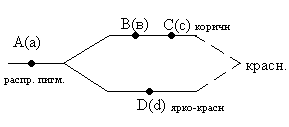 Р: ВВСС+ААdd aaDD F: ВВСС+АаDd F2: BBCC+
9:3:4 FA:АаDd aadd Расщепление 1:2:1 Р: ААВВ+ССdd ccDD F: ААВВ+СсDd F2: ААВВ+
9:3:3:1 в анализирующем скрещивании расщепление 1:1:1:1 При наследовании окраски глаз у дрозофилы возможны варианты расщепления 9:7, в анализирующем скрещивании 3:1. существует еще один вариант расщепления при комплиментарности 9:6:1, доказывается при изучении наследования формы плода у тыквы.
Доказательство:В анализирующем 1:2:1
Рецессивный эпистаз может трактоваться и как эпистаз и как комплиментарность. Полимерия – это такой тип взаимодействия неаллельных генов, при котором действие доминантного аллеля одного гена полностью или частично дублирует действие доминантного аллеля другого гена), есть на проявление признака необходимы доминантные аллели одного из взаимодействующих генов). Существует два варианта полимерии:
классическим примером некумулятивной полимерии является наследование махноногости у лошадей (кур).
Однако реально в данном случае расщепление 9:6:1. Некумулятивную полимерию можно трактовать как комплиментарность. По данному типу взаимодействия неаллельных генов наследуется форма плодов пастушьей сумки. В анализирующем скрещивании расщепление 1:2:1. Кумулятивная полимерия связана с неполным дублировании генов с неполным доминировании по каждому гену. Классическим примером является степень пигментации кожи. Реально степень пигментации зависит от 5 генов, однако для простоты будем считать, что она зависит только от 2 генов.
В анализирующем скрещивании расщепление 1:2:1 |