лекции по генетике. Лекция Законы наследования
 Скачать 211.76 Kb. Скачать 211.76 Kb.
|

|
Ген и его свойства.
Роль гена в биосинтезе белка заключается в том, что в процессе транскрибции считываются РНК, необходимые для образования всех компонентов белоксинтезирующего аппарата. В процессе транскрибции помимо различных функциональных белков считываются регуляторные белки, которые способны активировать или ингибировать действие структурных генов, т.к. гены эукориот имеют мозаичное строение, а матричные РНК интрон-экзонную структуру, то синтез различных белков и их активность регулируется за счет альтернативного сплайсинга. Генотип, как целостная система.
цитоплазматическое наследование. Геном включает в себя: совокупность ядерных генов или нуклиом, а также совокупность генов митохондрий и пластид. Совокупность митохондриальных генов, получило название хондриом. Совокупность генов пластид называется пластидом. Гены, локализованные в митохондриях и пластидах, называется плазмогенами, и существуют определенные закономерности их наследования, которые получили название цитоплазматическое наследование. Расхождение митохондрий и пластид в мейозе случайно, поэтому наследование плазмогенов не подчиняется законам Менделя и Моргана. Определенные закономерности наследования плазмогенов проявляется на фоне анизогамии. Во варианте анизогамии половые клетки отличаются друг от друга по количеству митохондрий и пластид, а следовательно, плазмогенов в сперматозоидах этих органов практически нет, кроме того при попадании в яйцеклетку митохондрий и пластид часто разрушаются, поэтому для будущего зародыша имеет значение только плазмогены матери. Такой вариант цитоплазматического наследования получил название матрокления. Классическим примером является наследование пестролистности у растений (например, герань). ♂пестролист ♀зеленые растения ↓ зеленый ♀пестролист ♂зеленые листья ↓ зеленые, пестрые, белые появление во втором случае разных листьев у герани объясняется с точки зрения явления митотической сегрегации. 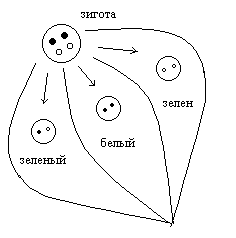 Для человека также характерно явление матроклении. В митохондриях человека содержится 36 генов, 22 для тРНК, 2 гена для собственных рРНК, а все остальное для белков цепи переносчиков, следовательно, при потологии этих генов могут наблюдаться дифекты окислительного фосфолилирования, энергообмен сдвигается к гликолизу и развивается лактоацидоз. Наблюдается митохондриальные болезни различной степени тяжести. 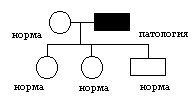 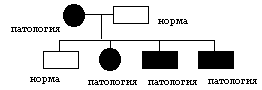 различная тяжесть заболевания является причиной митатической сигригации. Существуют определенные варианты псевдоматроклинии, например, у отрядя легочных брюхоногих моллюсков бывает левозакрученная и правозакрученная раковины. Оказалось, что закрученность раковины является результатом спирального дробления зиготы на ранних этапах развития зародыша. В это время гены зиготы не работают и правозакрученность раковины определяется только наличием белкка в цитоплазме зиготы, если данного белка нет, то раковина закручена влево. Наличие белка определяется доминантным аутосомным геном. Фенотип зависит от генотипа и это отражает явление изменчивости. Изменчивость. Это свойство живых систем и их компонентов существовать в различных вариантах. Выделяют следующии варианты изменчивости:
Лекция 5. Генотипическая изменчивость Определение смотри выше. Делится на два крупных варианта:
определение кроссинговера, история открытия (анализирующие скрещивание на дрозофилах) рисунок метофазы 1, объяснить вероятностный характер оплодотворения.
Существует много классификаций мутаций:
Генные мутации. Изменение структуры гена – это изменение числа нуклеотидов, либо изменение последовательности нуклеотидов. Такие мутации могут возникать спонтанно или могут быть вызваны ошибками "трех R ": репликация, рекомбинация (кроссинговер, произвольная ориентация бивалентов в метофазе 1), репарация. Наименьшие изменения структуры гена касаются изменения только в одной паре нуклеотидов, так называемые точковые мутации. При мутациях пуриновые нуклеотиды могут менятся на пуриновые, а перемидиновые на перемединовые, такие мутации называются транзиции. Входе мутации меняются пуриновые на перемединовые и наоборот такие муьтации называются трансверзии. Возможно три варианта точковых мутаций:
последствия замены: не происходит изменения в порлипептидной цепи (вырожденность генетического кода). Такие мутации не ведут к изменению жизнеспособности и называются сейнсис мутации. Может измениться первичная структура белка на одну аминокислоту (спецефичность генетического кода). Последствия такой замены различны, т.е. это либо миссенс мутации, когда белок функционирует плохо. Первичная структура белка может стать короче из-за появления нонсенс кодона, нонсенс мутации, прекращается синтез белка. Белок может удлиниться, если замена произойдет в нонсенс кодане. Белок может стать короче из-за изменения кадона инициации. Последствия вставки или выпадения более серьезны: Меняется структура полипептида с места выпадения или вставки (неперекрываемость и непрерывность генетического кода) из-за сдвина рамки считывания С большой вероятностью появляется нонсенс кодон и синтез белка прекращается. Генные мутации являются одной из причин возникновения новых аллелей и поэтому являются причиной множественного аллелизма. С этой точки зрения они бывают доминантными или рецессивными в ряде случаев генные мутации ведут к фенотипическим изменениям и к появлению различных наследственных заболеваний (полидактилия, дальтонизм, серповидно клеточная анемия, различные формы сахарного диабета, витамин Д независимый рахит). Спонтанные генные мутации являются ошибками "трех R ". Существуют индуцируемые мутации, которые вызваны действиями физикохимичеких агентов на ДНК. Частота спонтанных мутаций 10-6, индуцируемых 10-3 (тяжелые металлы, ультрафиолет, подавляющие большинство лекарственных препаратов). Если такие мутации произошли в генеративных клетках, то они передаются потомству, если в соматических , то возможно формирование раковой опухоли. Поэтому большинство мутагенов является канцерогенов. В клетках существует механизмы исправления генетических повреждений и восстановления нормальной структуры ДНК. Такие механизмы получили название репарации. Репарации. Существуют несколько вариантов репарации:
темновая репарация происходит в темноте и связана с вырезанием патологичного участка ДНК ферментом экзонуклеазой, затем брешь достраивается β-ДНК-полимеразой по правилу комплиментарности, окончательное сшивание цепи ДНК осуществляется лигазой, это ферментативный процесс, который идет с затратой АТФ. У человека существуют патологии при которых данный вид репарации не работает, в результате возникает заболевание пигментная ксеродерма (вид рака кожи). Пострепликативная проходит в том случае, если не срабатывают механизмы темновой репорации, проходит после репликации, когда синтезируется патологичная ДНК. 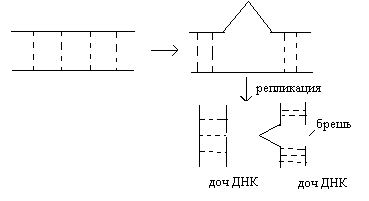 Если не включается механизм репарации, то патологичные ДНК расщепляются. Механизмы пострепликативной репарации основаны на внутригенном кроссинговере. 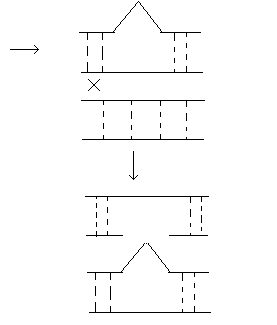 В первой молекуле по нашему рисунку брешь достраивается β-ДНК-полимеразой и включается лигаза. Вторая молекула подвергается темновой репорации. Если патологична пострепликативная репарация, то возникает пигментная ксеродерма. Хромосомные мутации. Это изменения структуры хромосом. Выделяют два варианта:
К внутрихромосомным мутациям относятся:
Делеция или нехватка заключается в выпадении какого-либо участка хромосом. Дуплекация – это удвоение какого-либо участка ДНК, возможны триплекации, тетропликации и т.д. Инверсия – это поворот участка хромосом на 1800. Транспозиция – это перемещение участка хромосом. Образование кольцевых хромосом – в этом случае слипаются концы хромосом, образуя кольцо. При репликации возможно разрушение кольцевых хромосом. Последствия делеции и дупликации. наиболее опасны последствия делеции. Причем важно какие гены выпали, домашнего хозяйства или гены роскоши. Последствия при делеции как правило несовместимы с жизнью. У гетерозигот по делеции наблюдаются тяжелые последствия связанные с нарушением баланса генов. У дрозофилы существует делеция "ночь", которая заключается в возникновении вырезки на крыле. У человека известна дилеция по пятой паре хромосом, которое происходит в коротком плече хромосомы. отставание в физическом и умственном развитие погибают в первые годы жизни. характерно изменение гортани, при котором новорожденный издает специфический крик, синдром кошачьего крика. Последствия дупликации обычно более мягкие, чем последствия дилеции. Однако нарушается баланс генов, что отражается на фенотипе. У дрозофилы известна дуплекация Bar , которая приводит к уменьшению фасеток в глазах. у гетерозигот число фасеток в глазах небольшое, у гомозигот число фасеток резко уменьшается, возникает полосковидный глаз. Механизмы делеции и дупликации. До недавнего времени считалось, что данные мутации возникают из-за разрыва ДНК, а затем из-за ее дальнейшего сшивания. В настоящее время считают, что это возможно, но только под действием очень сильных мутагенов. Основной причиной спонтанного возникновения дилеции и дупликации является кроссинговер. В ДНК очень много гомологичных последовательностей, которые могут конъюгировать между собой. Из-за неравноценного кроссинговера в гомологичных хромосомах возникает делеция или дуплекация. 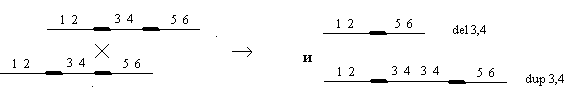 Последствия инверсии. Последствия наблюдаются как в гомозиготах, так и в гетерозиготах, однако инверсия приводит только к изменению порядка в расположении генов, поэтому они могут вообще не оказывать влияние на фенотип. У гомозигот по инверсии последствия наблюдаются редко, так как между гомологичными хромосомами может проходить нормальный кроссинговер. У гетерозигот по инверсии наблюдается высокий процент стерильности из-за образования несбалансированных гамет и высокий процент возникновения патологий у потомства. Это происходит из-за того, что гомологичные хромосомы не могут нормально конъюгировать и при конъюгации образуется так называемая инверсионная петля. 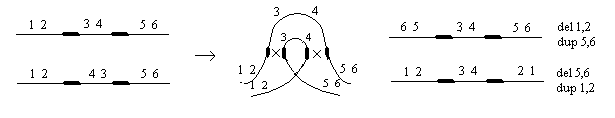 Инверсии являются эволюционным механизмом, который создает изоляцию между многими близкими видами, так как для близких видов часто характерны одинаковые инверсии. Механизм образования инверсии. Гомологичные участки ДНК в пределах одной молекул могут конъюгировать между собой. 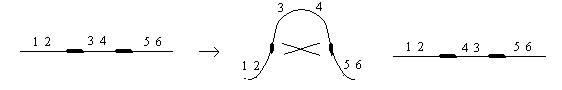 Транспозиции. При транспозиции все гены остаются на месте, но часть из них меняют свое положение. Последствий может не наблюдаться однако, механизм образования транспозиций связан с образованием кольцевых хромосом при разрыве такой кольцевой хромосомы происходит потеря генов и, следовательно, последствия будут аналогичны дилеции. 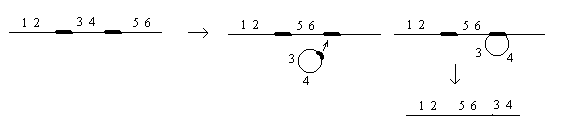 Образование кольцевых хромосом. Механизм смотри выше. При репликации кольцевых хромосом они характеризуются очень высоким уровнем нестабильности. При репликации кольцевых хромосом часто наблюдается перекручивание ДНК, когда кольцо попадает в кольцо. Тогда при расхождении кольцевых ДНК будет наблюдаться разрыв молекул и утраты генов. Последствия будут сходны с последствиями дилеции. Межхромосомные мутации. Этот вид мутации получил название транслокации. При транслокации происходит обмен участками ДНК входе кроссинговера между негомологичными хромосомами. Это возможно благодаря тому, что в ДНК негомологичных хромосом существуют короткие гомологичные последовательности. 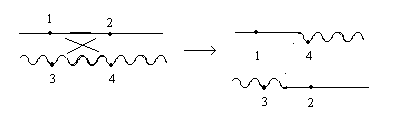 существуют как гомо-, так и гетерозиготы по транслокации. У гомозигот как правило последствий не наблюдается, т.к. все гены остаются на месте. У гетергзигот по транслокации, как правило, наблюдается высокий процент стерильности в потомстве и аномалии. Это связано с тем, что не гомологичные хромосомы в мейозе выстраиваются в форме креста. 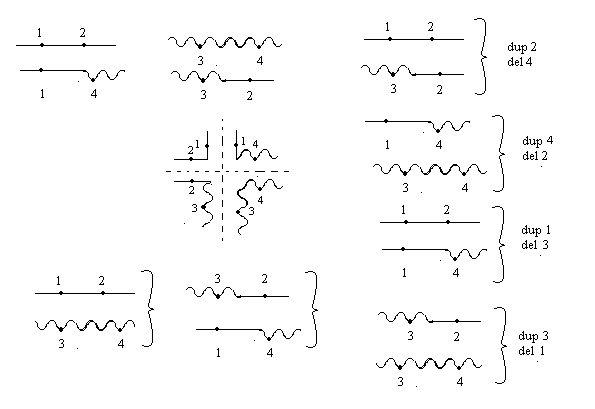 Геномные мутации. Это мутации связанные с изменением количества отдельных хромосом, либо с изменением целых хромосомальных наборов. Делятся на два варианта:
|