Нервная система. Нервная система (1). Нервной системы и высшей нервной деятельности
 Скачать 2.09 Mb. Скачать 2.09 Mb.
|

|
ТЕМА 5: Механизм проведение возбуждения по нервному волокну. ВОПРОСЫ: Строение нервного волокна. Механизм возникновения ПД в нервном волокне. Законы проведения возбуждения в нервном волокне ОСНОВНЫЕ ПОЛОЖЕНИЯ ТЕМЫ По строению нервные волокна бывают мякотные и безмякотные. Мякотные имеют осевой цилиндр и покрыты 2-мя оболочками – миелиновой и швановской. Миелин покрывает невное волокно участками. Пространство между миелином называют «перехваты Ранвье». В них формируются импульсы возбуждения. Миелин обладает большим сопротивлением, и поэтому передача возбуждения по мякотному волокну осуществляется скачкообразно с большой скоростью (Рис.21). Швановская оболочка обладает трофической функцией и большим сопротивлением, выполняя роль изолятора. 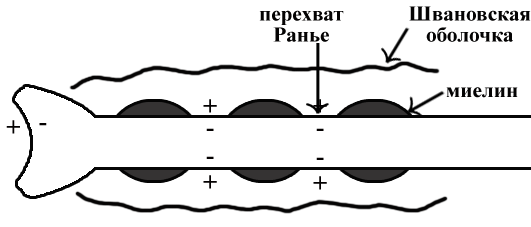 Рис.21. Строение мякотного нервного волокна. Безмякотные волокна не имеют миелина, поэтому возбуждение постепенно охватывает каждый соседний участок (Рис.22). Скорость передачи нервного импульса в таком волокне меньше, чем в мякотном. 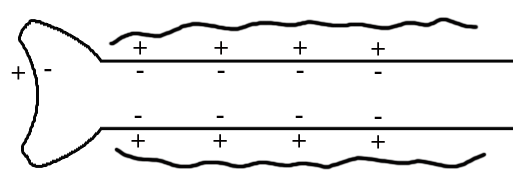 Рис.22. Строение безмякотного нервного волокна. По скорости передачи возбуждения нервные волокна делятся на 3 группы: Группа А. Скорость проведения 70 – 120 м/с. Это волокна чувствительные, двигательные. Группа В. Скорость проведения 15 м/с. Это волокна чувствительные и преганглионарные вегетативные. Группа С. Скорость проведения 5м/с. Это постганглионарные вегетативные нервные волокна, исходящие из вегетативных ганглиев, где нет миелина. Механизм проведения возбуждения в нервном волокне При действии раздражителя в рецепторной клетке возникает рецепторный потенциал (РП.), который является локальным током и по ткани не распространяется. В случае вторичного рецептора под влиянием РП из клетки выделяется ацетилхолин, который действует на окончание чувствительного нервного волокна, увеличивает в мембране проницаемость для ионов натрия , вызывая частичную деполяризацию, которая называется генераторным потенциалом (ГП). Генераторный потенциал является копией рецепторного потенциала. Если раздражитель действует прямо на нервное окончание в случае первичного рецептора, то в нём возникает рецепторный потенциал (РП). Частичная деполяризация, возникшая в окончании нервного волокна (РП или ГП), вызывает транспорт ионов натрия в сторону « - « заряда до уравнивания концентраций. В цитоплазме ионы натрия перемещаются от окончания к первому перехвату Ранвье, а над мембраной в обратном направлении. Этот горизонтальный переток ионов натрия обеспечит частичную деполяризацию в первом перехвате Ранвье. Если она достигает критического уровня – возникает потенциал действия (Рис.23). 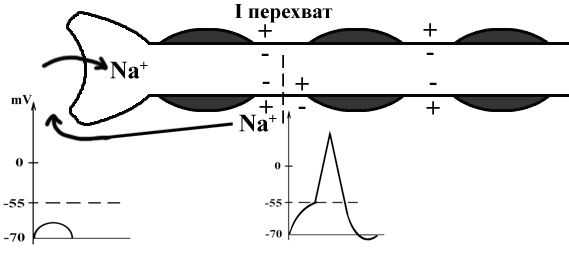 Рис.23. Механизм возникновения потенциала действия в нервном волокне. В первом перехвате Ранвье возникла полная деполяризация с изменением зарядов ( в цитоплазме “+” за счёт скопления ионов натрия, а над мембраной “-‘ за счёт потери натрия). Далее ионы натрия, перемещаясь в цитоплазме в сторону “- “ заряда, будут формировать потенциал действия (ПД) во втором перехвате Ранвье ( Рис.24). 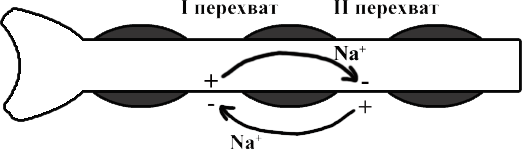 Рис. 24. Механизм передачи возбуждения в мякотном волокне. В безмякотном волокне нет изолятора миелина, поэтому транспорт ионов натрия будет осуществляться между соседними участками (Рис.25). 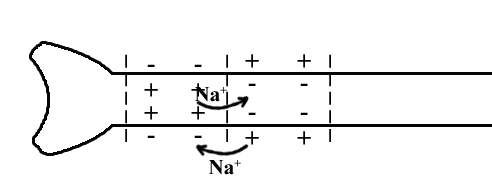 Рис. 25. Механизм передачи возбуждения в безмякотном волокне. Как только мебранный потенциал во втором участке упадёт до -55 мв., в нём возникает потенциал действия (ПД), а в первом участке – абсолютная рефрактерность (Рис. 20). ТЕМА 6: Физиология нервного центра ВОПРОСЫ: Строение синапса Механизм передачи возбуждения в нервном центре. Свойства нервного центра ОСНОВНЫЕ ПОЛОЖЕНИЯ ТЕМЫ Нервный центр – это совокупность нейронов, расположенных на различных этажах ЦНС и регулирующих деятельность определённого органа. В нервном центре выделяют рабочий отдел и надстройку. Надстройка представлена нейронами, расположенными в выше лежащих отделах ЦНС и управляющими деятельность рабочего отдела. Например: центр ССС деятельности состоит из нейронов, расположенных на уровне продолговатого мозга, гипоталамуса и коры головного мозга. Рабочим отделом являются нейроны в области продолговатого мозга. Нейроны в области гипоталамуса и коры головного мозга входят в состав надстройки. Структурной частью нервного центра является нейрон (Рис.26). Он имеет тело неправильной формы и 2 вида отростков: многочисленные короткие дендриты и одиночный длинный отросток – аксон. По дендриту возбуждение идёт к телу нейрона. По аксону – от нейрона к рабочему органу или другому нейрону. 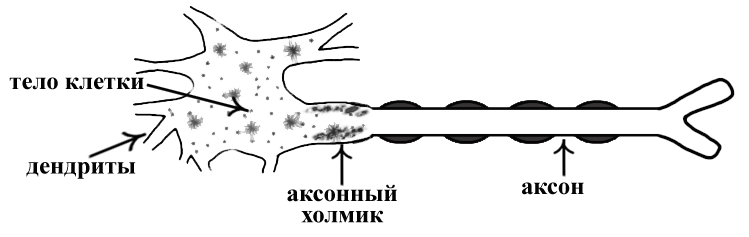 Рис.26. Строение нейрона По функции нейроны делятся на: Чувствительные – афферентные, расположенные в спинномозговых ганглиях, ядрах черепных нервов, спинном и головном мозге. Двигательные – эфферентные, находящиеся в коре, подкоркой области, стволе головного мозга, передних рогах спинного мозга. Ассоциативные – вставочные. Объединяющие, передающие импульсы с афферентного на афферентные нейроны. Нейросекреторные ( например в гипоталамусе). Обладающие свойством вырабатывать и выделять в кровь гормоны, названные нейросекретами. Место контакта двух нейронов называется СИНАПС. Его формируют либо разветвление аксона и тело нейрона, либо аксон и дендрит. Строение синапса (Рис.27): А. Пресинаптичекая мембрана ( мембрана, покрывающая окончание аксона в месте контакта).Пресинаптические окончания образуют бляшки, в которых находятся везикулы, содержащие медиатор. С помощью медиатора возбуждение передаётся через синаптическую щель на постсинаптическую мембрану. Б. Синаптическая щель (расстояние между пре и постсинаптической мембранами 20 – 40 н/м.). В период возбуждения в синаптической щели появляются адгезивные белки, которые фиксируют положение пре и постсинаптических мембран и способствуют точной передачи медиатора. В. Постсинаптическая мембрана (часть мембраны нейрона, содержащая рецепторы к медиатору). Медиаторы могут быть различными и возбуждающими и тормозными. Постсинаптическая мембрана является частью постсинаптической плотности, куда входят 1000-1500 белков (арматурные белки, белки цитоскелета, актомиозин). 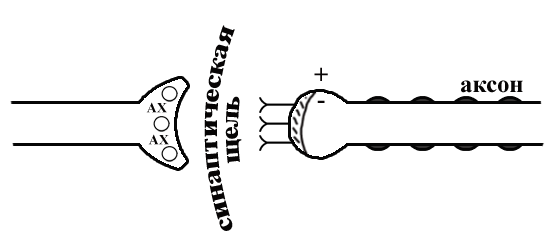 Рис.27. Структура синапса. Механизм передачи возбуждения в синапсе (на примере медиатора ацетилхолина). Импульс возбуждения, подошедший к пресинаптической мембране, увеличивает её проницаемость для ионов кальция, который входит в синаптическую бляшку, связывается с белком и возникает выброс медиатора из бляшки. Квант медиатора проходит через синаптическую щель, контактирует с рецетором постсинаптической мембраны, увеличивает частично её проницаемость для ионов натрия и вызывает частичную деполяризацию на 3-5 мв. (возбуждающий постсинаптический потенциал или ВПСП). Чтобы получить возбуждение в нервном центре, необходимо суммировать 3-5 таких ВПСП для достижения критического уровня деполяризации (Рис.28). Для этого необходимо к пресинаптической мембране подвести минимум три импульса возбуждения и выбить 3 кванта медиатора. Потенциал действия возникает в аксонном холмике нейрона. 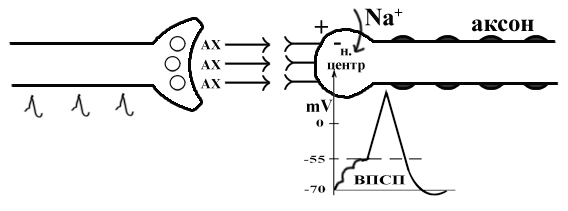 Рис.28. Генерация потенциала действия в нейроне. Генерация потенциала действия в нейроне прекращается ретрограднеой сигнализацией (Рис.29). Во время возбуждения нейрона активируются дополнительно метаботропные рецепторы, которые увеличивают проницаемость постсинаптической мембраны для ионов кальция. Кальций, попав в цитоплазму нейрона, активирует фосфолипазу, которая выщепляет арахидоновую кислоту из мембраны. Из неё образуется медиатор 2-АГ 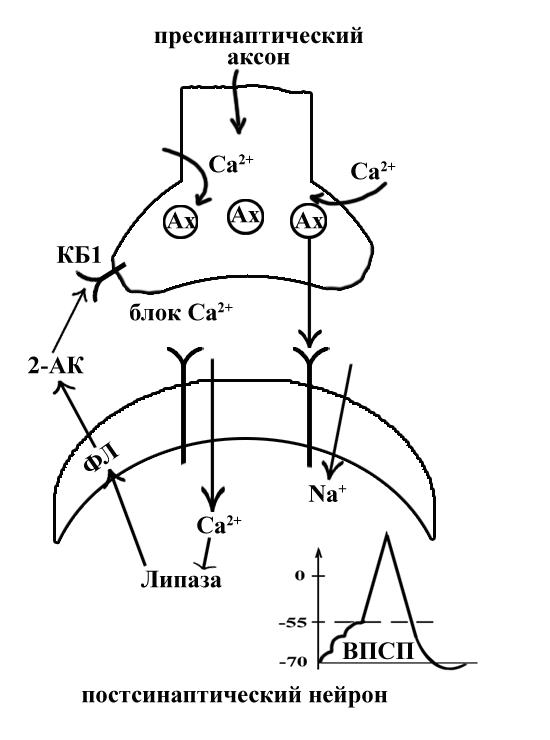 Рис. 29. Схема ретроградной сигнализации в синапсе (2-арахидоноилглицерин), который транспортируется через синаптическую щель назад к пресинаптической мембране, находит для себя рецептор КБ-1 (канабиоидные). Возбуждение этих рецепторов блокирует ток кальция в пресинаптической мембране и медиатор из везикул не выделяется. Генерация потенциала действия в нейроне прекращается. Таким механизмом регуляруется частота нервных импульсов в нейроне. СВОЙСТВА НЕРВНОГО ЦЕНТРА Односторонняя передача возбуждения в синапсе ( от пре к постсинаптической мембране ). В обратном направлении передача невозможна, ибо медиатор содержится только в пресинаптических бляшках, а рецептор к нему только на постсинаптической мембране. Синаптическая задержка. В синапсе передача возбуждения задерживается на 0,6 – 0,8 сигм ( 1 сигма=0,001 сек.). Время уходит на выделение медиатора, транспорт его через синаптическую щель, контакт с рецептором и суммацию ВПСП. Суммация ВПСП. А. Последовательная, когда последователь по времени суммируются ВПСП. Б. Пространственная (одновременная). На одном нейроне может контактировать несколько аксонов. Из каждого одновременно выделится по кванту АХ, которые сразу вызовут падение мембранного потенциала в нейроне до критического уровня (Рис.30). 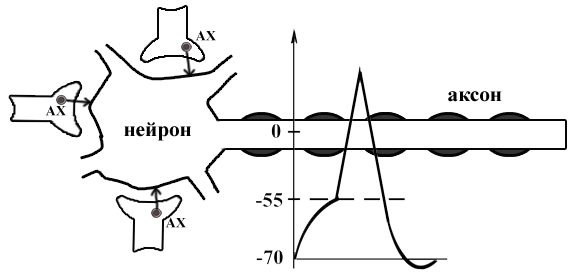 Рис. 30. Схема одновременной суммации ВПСП. Высокая утомляемость нервного центра. При длительном действии раздражителя расходуется АХ в пресинаптической области и не возбуждается постсинаптическая мембрана. Восстанавливается работоспособность утомлённого нервного центра за счёт отдыха. Он может быть активным и пассивным. Пассивный отдых. Когда ничего не делая ожидается время синтеза достаточного количества АХ в пресинаптической бляшке. Активный отдых. Для этого необходимо заняться другим видом работы и подключить к возбуждению параллельную рефлекторную дугу. От неё по коллатерали возбуждение подойдёт к утомлённому нервному центру и выбросит в синапс недостающее количество АХ. ВПСП одновременно будет суммироваться на нейроне из нескольких синапсов. Этот вид отдыха более продуктивен, восстановление трудоспособности наступает быстрее, чем при пассивном отдыхе (Рис.31). 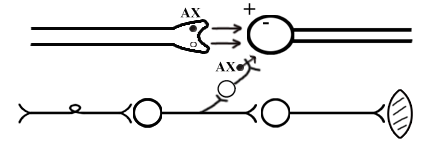 Рис. 31. Механизм активного отдыха. ТРАНСФОРМАЦИЯ ритма. Нервный центр может изменять число нервных импульсов, подошедших к нему в область пресинаптичеческих бляшек, увеличивая или уменьшая частоту (трасформируя). Поэтому трансформация может быть как поышающая, так и понижающая. ПОНИЖАЮЩАЯ – возникает за счёт суммации ВПСП (Рис.32). 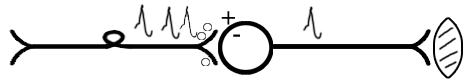 Рис.32. Механизм понижающей трансформации. ПОВЫШАЮЩАЯ трансформация за счёт возвратных коллатералей, исходящих от нервного центра. Из окончаний собственных коллатералей выделяется АХ, который вызовет дополнительные импульсы возбуждения в нервном центре ( Рис.33). 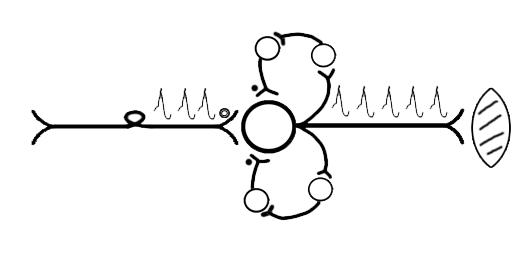 Рис. 33. Механизм повышающей трансформации в нервном центре. ПОСЛЕДЕЙСТВИЕ – способность нервного центра генерировать возбуждение после действия раздражителя. Это может быть связано с действием гуморальных раздражителей, способных вызывать деполяризацию или с наличием возвратных коллатералей. По ним импульсы возвращаются к нейрону и его возбуждают (Рис.34). 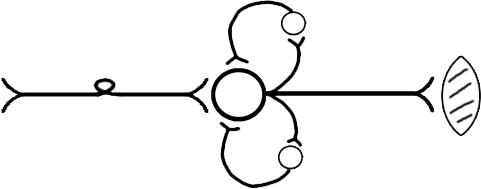 Рис.34. Механизм последействия в нервном центре. ВЫСОКАЯ ЧУВСТВИТЕЛЬНОСТЬ нервного центра к биологически активным веществам (БАВ). В крови существуют соединения, способные увеличивать проницаемость мембраны нейрона к ионам натрия, вызывая деполяризацию. При небольшой их концентрации возникает частичная деполяризация, что определяет тонус нейрона и его готовность к ответной реакции. Это важно для поддержания гомеостаза в организме. При высокой концентрации БАВ в крови может возникнуть самовозбуждение нейронов без действия раздражителей. Нервный центр обладает высокой чувствительностью к недостатку кислорода. Нейроны коры головного мозга способны существовать без кислорода не более 3-5 минут и этим определяется длительность клинической смерти. Нейроны нижележащих отделов ЦНС могут существовать без кислорода несколько дольше. НИЗКАЯ ЛАБИЛЬНОСТЬ нервного центра. Лабильность – это функциональная активность. К нервному центру подходит до 500 импульсов, а он может пропустить 100-120 импульсов. Это связано с последовательной суммацией ВПСП, когда частота импульсов теряется. Низкая лабильность предохраняет рабочий орган от перегрузок. ТЕМА 7: Торможение в центральной нервной системе. ВОПРОСЫ: Пресинаптическое торможение. Постсинаптическоеторможение. Надсегментарное торможение. ОСНОВНЫЕ ПОЛОЖЕНИЯ ТЕМЫ ТОРМОЖЕНИЕ – одно из свойств нервного центра, но рассматривается отдельно из-за сложности в организации. По механизму возникновения торможение делится на 2 вида - пре и постсинаптическое. ПРЕСИНАПТИЧЕСКОЕ возникает на пресинаптической мембране. Существует Д-нейрон, который связывает две параллельные рефлекторные дуги через коллатераль. Он контактирует с пресинаптической мембраной. Медиаторы, выделяющиеся в этих тормозных синапсах, вызывают стойкую деполяризацию пресинаптических структур. Возникшая деполяризация сопровождается длительной абсолютной рефрактерностью, которая обуславливает частичную или полную блокаду проведения нервного импульса к центру. ПОСТСИНАПТИЧЕСКОЕ торможение возникает на постсинаптической мембране при действии на неё ГАМК (гамма-аминомасляная кислота). Увеличивается проницаемость постсинаптической мембраны для ионов хлора и калия, возникает гиперполяризация в нейроне. При одном кванте, выделяющейся ГАМК в нейроне возникает гиперполяризация + 3-5 мв и называется ТПСП – тормозный постсинаптичекий потенциал. Чем больше выделяется ГАМК - тем больше гиперполяризация, которая увеличивает порог деполяризации и уменьшает возбудимость (Рис.35). 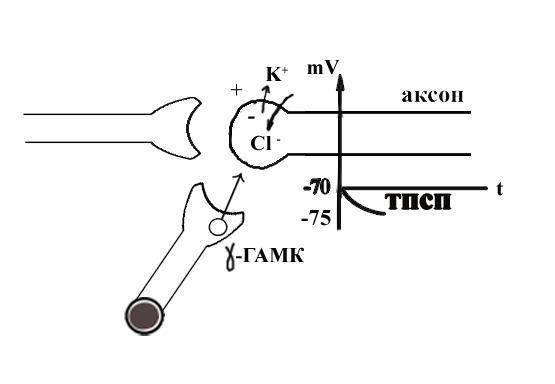 Рис. 35. Механизм постсинаптического торможения. ВОЗВРАТНОЕ торможение – это вариант постсинаптического торможения, которое осуществляется через клетку Рэншоу (Рис.36). При большой частоте нервных импульсов поступающих к нейрону, он перебрасывает часть импульсов через собственную коллатераль тормозной клетке Рэншоу. Она выделяет ГАМК и тормозит нейрон за счёт гиперполяризации. Это торможение охранительное, оно охраняет нейрон от перегрузок. 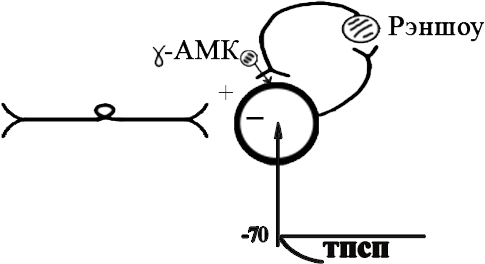 Рис. 36. Механизм возвратного торможения. По уровню расположения тормозного нейрона торможение делится на сегментарное и надсегментарное. СЕГМЕНТАРНОЕ. Тормозный нейрон находится на уровне одного сегмента с возбуждающим нейроном и может его тормозить по механизму либо пре, либо постсинаптичекого торможения. Тормозные нейроны в физиологических схемах обозначаются заштрихованными (Рис.37). 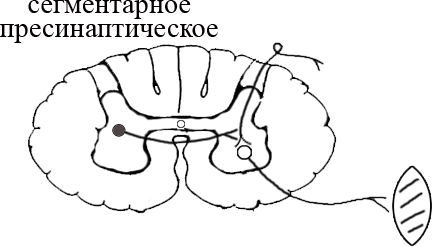 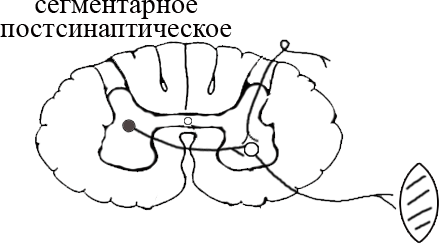 Рис.37. Варианты сегментарного торможения. НАДСЕГМЕНТАРНОЕ торможение открыл наш великий русский физиолог Иван Михайлович Сеченов (Рис.38). Он опустил лапку лягушки в 0,1н раствор серной кислоты, и она тотчас её выдернула (рефлекс Тюрка). У этой лягушки вскрыли черепную коробку и на зрительные бугры наложили кусочек соли. Теперь лапка свободно находилась в растворе серной кислоты. Далее смыли соль с области зрительных бугров и рефлекс Тюрка воздействие слабого раствора серной кислоты у лягушки восстановился. 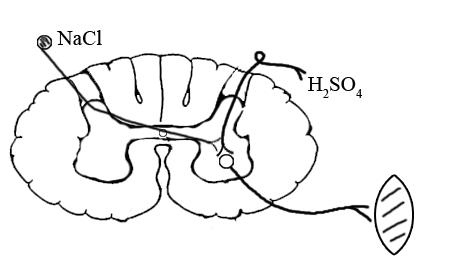 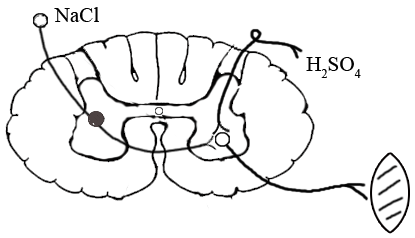 Рис. 38. Варианты надсегментарного торможения. И.М.Сеченов сделал вывод о том, что в зрительных буграх соль вызвала возбуждение, которое затормозило рефлекс Тюрка. Он назвал это возбуждение «тормозящим». Сейчас доказано, что в центральной нервной системе есть специальные тормозные нейроны, которое могут вызвать торможение в любых отделах, включая спинной мозг. |