Нервная система. Нервная система (1). Нервной системы и высшей нервной деятельности
 Скачать 2.09 Mb. Скачать 2.09 Mb.
|

|
ТЕМА 3: СТРОЕНИЕ МЫШЦЫ И МЕХАНИЗМ МЫШЕЧНОГО СОКРАЩЕНИЯ. ВОПРОСЫ: Строение мышцы, саркомера. Механизм мышечного сокращения. Динамика возбудимости в процессе возбуждения. Фазы мышечного сокращения. Зубчатый тетанус. Гладкий тетанус. Оптимум и пессимум частоты. ОСНОВНЫЕ ПОЛОЖЕНИЯ ТЕМЫ Все мышцы у позвоночных и у человека делятся на 2 типа: Гладкая мускулатура, которая имеется во внутренних органах и стенках сосудов. Поперечнополосатая – а) сердечная, б) скелетная Скелетная (поперечнополосатая) мускулатура выполняет следующие функции: перемещение тела в пространстве перемещение частей тела относительно друг друга поддержание позы Структурно-функциональной единицей поперечно-полосатой мышцы является нейромоторная единица (НМЕ). Она представлена аксоном мотонейрона, его разветвлениями и мышечными волокнами, которые иннервируются ими. Структура мышечного волокна Каждая мышца состоит из мышечных волокон, расположенных продольно, которые представляют собой многоядерные клетки. Снаружи они покрыты базальной мембраной и плазмолеммой, между которыми располагаются камбиальные клетки (миосателлиоциты). На плазмолемме во многих местах имеются пальцеобразные вдавления – Т-тубулы. Они связывают сарколемму с саркоплазматическим ретикулюмом (СПР). Внутри имеется обычный набор органелл: многочисленные ядра, занимающие периферическое положение, митохондрии и т.д. СПР – это система связанных между собой канальцев с высоким содержанием Ca+ Центральную часть цитоплазмы занимают специфические органеллы – миофибриллы – сократительные элементы, расположенные продольно. 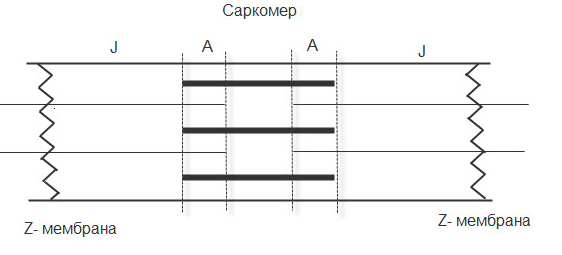 Рис.10. Строение саркомера Структурной единицей миофибрилл является саркомер. Это постоянно повторяющаяся часть миофибриллы, заключенная между двумя Z-мембранами (телофрагмами). Посредине саркомера имеется линия М – мезофрагма. К мезофрагме крепятся нити миозина – сократительного белка, а к телофрагме – актин (тоже сократительный белок). Чередование этих сократительных белков составляет поперечную исчерченность (Рис.10). В саркомере выделяют анизотропный диск (А) – диск с двойным лучепреломлением (миозин + концы актина), Н-зону – только нити миозина (входит в состав диска А) и I-диск – только нити актина. При сокращении саркомера происходит укорочение диска I и уменьшение светлой зоны Н. Сокращение всей мышцы определяется укорочением саркомера, а его длина сокращается за счет образования акто-миозиновых комплексов. Миозин – толстая белковая молекула, которая располагается по ценру саркомера и состоит из двух цепей – легкого и тяжелого меромиозина. На поперечном сечении миозин имеет вид ромашки – центральная часть и отвисающие головки. Головка легкого меромиозина обладает АТФ-азной активностью, которая проявляется лишь в момент контакта с активным участком актина. 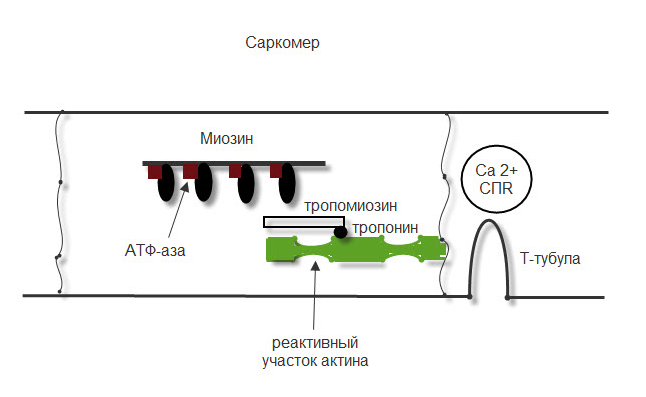 Рис.11. Расположение сократительных белков в саркомере. Актин – глобулярный белок, состоит из двух цепей, переплетенных между собой в виде бус. На каждой глобуле имеются активные участки, которые закрыты тропомиозином, и его положение регулируется тропонином. В состоянии покоя активные участки актина не взаимодействуют с головкой миозина, так как они прикрыты в виде крышки тропомиозином (Рис.11). Механизм мышечного сокращения. При возбуждении мотонейрона импульсы подходят к мионевральной пластинке (место контакта аксона и плазмолеммы). Из пресинаптической мембраны выделяется ацетилхолин (АХ), который проходит синаптическую щель и действует на плазмолемму (в этом месте ее можно назвать постсинаптической), находит рецепторы к АХ и взаимодействие с ними отражается на проницаемости мембраны для ионов натрия. Проницаемость мембраны для натрия повышается, возникает деполяризация, что приводит к возникновению ПД. Он распространяется вдоль мембраны и передается на Т-тубулы, которые тесно связаны с СПР. ПД в области Т-тубул вызывает повышение проницаемости мембраны СПР для кальция, и он выбрасывается в цитоплазму квантами (порциями) в зависимости от частоты импульса. Кальций запускает механизм укорочения саркомера. От концентрации кальция зависит насколько сократится саркомер (и мышца в целом). Выброшенный в цитоплазму кальций находит белок тропонин, взаимодействует с ним и вызывает его конформационные изменения (то есть меняет пространственное расположение белка). Конформационные изменения тропонина сдвигают тропомиозин со своего места, при этом открывается активный (реактивный) участок актина. В этот открытый участок встраивается головка миозина. При этом контакте активируются ферментативные системы, расположенные последовательно. И этот контакт двух белков по типу зубчатой передачи механически передвигает нить актина к центру саркомера. Возникает актиновый шаг. Чем больше возникает актиновых шагов, тем сильнее укорачивается саркомер. В момент контакта головки миозина и реактивного участка актина головка приобретает АТФ-азную активность. На что расходуется энергия АТФ: - на гребкообразное движение и разрыв связей между актином и миозином; - на работу кальциевого насоса; - на работу натрий-калиевого насоса. Таким образом, чем больше выделяется кальция, тем больше образуется акто-миозиновых комплексов, тем больше гребков делает миозин, тем сильнее укорачивается саркомер. Как только мотонейрон перестает посылать импульсы к мембране мышечного волокна, и в СПР перестает поступать ПД от Т-тубул, выброс кальция из СПР прекращается, и усиливается работа кальциевого насоса, разрываются акто-миозиновые мостики, Z-мембрана возвращается на место и происходит расслабление саркомера (и мышцы в целом). Фазы мышечного сокращения. Мышечное сокращение можно зарегистрировать на кимографе. Для этого мышца крепится к штативу, а к другому концу – писчик, который записывает мышечное сокращение (Рис.12). 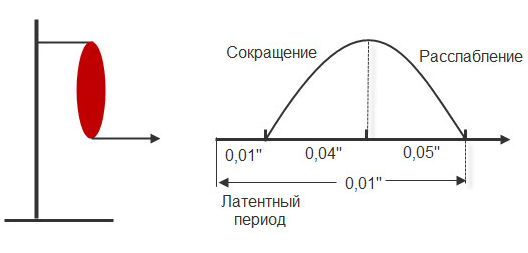 Рис.12. График одиночного мышечного сокращения. В мышечном сокращении выделяют следующие фазы: - латентная (0,01 сек) – от начала действия раздражителя до видимой ответной реакции; - фаза сокращения (0,04 сек); - фаза расслабления (0,05 сек). Таким образом, одиночное мышечное сокращение занимает 0,1 сек. В период мышечного сокращения меняется возбудимость ткани, то есть ее способность к повторной ответной реакции при действии высокочастотных раздражителей. При относительно низких частотах ответная реакция будет выглядеть как серия одиночных мышечных сокращений (до 10 импульсов в секунду). Тетанусы. Оптимум и пессимум частоты. Если увеличить частоту наносимых раздражителей, то можно подобрать такую частоту, при которой каждый последующий раздражитель подействует в фазу расслабления. При этом мышца сократится из неполностью расслабленного состояния, и ответной реакцией будет зубчатый тетанус. Для икроножной мышцы лягушки зубчатый тетанус возникает при частоте больше 10, но меньше 20 импульсов (каждый последующий импульс приходит через 0,09 – 0,06 сек) При дальнейшем увеличении частоты более 20 импульсов в секунду (до 50) регистрируется гладкий тетанус, так как каждый импульс попадает в период сокращения, и мышца сокращается из сокращенного состояния (каждый последующий импульс приходит через 0,02 – 0,05 сек). 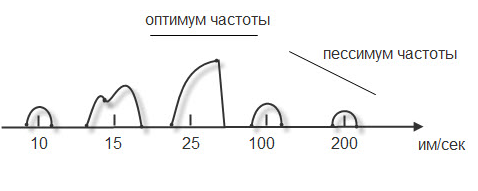 Рис.13. Характер ответной мышечной реакции при увеличении частоты наносимых раздражений. Зубчатый тетанус выше, чем одиночное мышечное сокращение, а гладкий еще выше. В основе тетануса лежит суммация (суперпозиция) сокращений и высокая концентрация кальция, выброшенного из СПР. При увеличении частоты раздражителя увеличивается выброс кальция из СПР, который выделяется квантами и не успевает вернуться обратно. Но не всякие высокочастотные раздражители вызывают оптимальное сокращение. Чаще всего оптимальное сокращение вызывает гладкий тетанус. Оптимум частоты – максимальная ответная реакция на действие высокочастотных раздражителей. Раздражители очень высокой частоты могут уменьшать ответную реакцию, и тогда возникает пессимум частоты. При частоте 100 импульсов в секунду раздражитель попадает в конец латентной фазы (каждый последующий импульс приходит через 0,01 сек), и в ответ возникает одиночное мышечное сокращение. При частоте 200 имп/сек (каждый последующий импульс приходит через 0,005 сек) возникает либо одиночное мышечное сокращение, либо реакция отсутствует. Уменьшение ответной реакции в период пессимума связано с действие раздражителя в период либо абсолютной, либо относительной рефрактерности. Абсолютная рефрактерность занимает 0,005 сек. Затем, в период относительной рефрактерности, возбудимость ниже 100%. Возбудимость восстанавливается через 0,01 сек. (Рис.13). ТЕМА 4: Строение рефлекторной дуги. Классификация и свойства рецепторов ВОПРОСЫ: Нейро-гуморальная регуляция. Что такое «рефлекс»? Строение рефлекторной дуги. Рецептивное поле рефлекса. Классификация рецепторов. Свойства рецепторов. ОСНОВНЫЕ ПОЛОЖЕНИЯ ТЕМЫ Каждый орган выполняет определённую функцию. Изменение функции одного органа влияет на работу других органов. Координирует работу всех органов нервная система. Посылая импульсы к каждому органу, нервная система иннервирует и железы внутренней секреции, которые выделяют гормоны. Последние через кровь изменяют функциональное состояние всех систем организма. Регуляция, осуществляемая биологически активными веществами, циркулирующими в крови, называется гуморальной. Таким образом, в организме существуют два пути регуляции: нервный и гуморальный. Эти пути имеют некоторые отличия: Нервная регуляция точная, т.к. нервные волокна идут к точному адресату. Гуморальная регуляция через кровь подключает к ответу множество органов. Нервная регуляция быстрая (скорость передачи нервного импульса 120 м/сек.). Гуморальная регуляция замедленная (скорость кровотока 0,5 м/сек.). Эффект нервной регуляции быстро заканчивается (мышца сократилась). Эффект гуморальной регуляции длится долго, до тех пор пока не инактивируется биологически активное соединение в кровотоке. Обе эти системы регуляции работают всегда вместе. Не существует в организме только нервной или только гуморальной регуляции . ФУНКЦИИ НЕРВНОЙ СИСТЕМЫ 1. Поддерживает гомеостаз (сохранение постоянства внутренней среды организма). Под этим понимается постоянство температуры, РН, осмотического и онкотического давления, газового состава, питательных веществ в крови, лимфе, тканевой жидкости и т.д. 2. Координирует деятельность отдельных органов. 3. Осуществляет взаимосвязь организма с внешней средой. Основной акт, который осуществляет нервная система – это рефлекс. РЕФЛЕКС – это ответная реакция организма, осуществляемая нервной системой при действии раздражителя. Морфологической основой рефлекса является РЕФЛЕКТОРНАЯ ДУГА. Это путь, по которому осуществляется рефлекторная деятельность. В каждой рефлекторной дуге выделяют 5 звеньев (Рис.15): Рецептор (рецепторная клетка или терминаль дентрита). Афферентный путь (чувствительный, центростремительный). Включает дендрит, чувствительную нервную клетку, её аксон, идущие к нервному центру. Нервный центр (эфферентная клетка). Нервный центр может усложняться за счёт вставочных нейронов. Эфферентный отдел (аксон от эфферентного нейрона). Эффектор (рабочий орган). 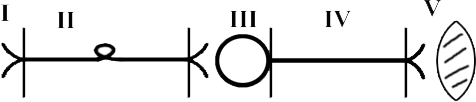 Рис.15. Схема рефлекторной дуги. По количеству нейронов рефлекторные дуги бывают: Двухнейронные (чувствительный и эфферентный нейрон). Многонейронные (много нейронов в нервном центре) (Рис.16). 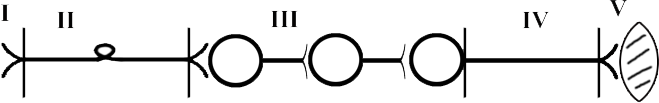 Рис.16. Схема многонейронной рефлекторной дуги. СТРОЕНИЕ РЕЦЕПТОРА Рецептором может являться специализированная клетка , контактирующая с терминалями нервного волокна. Оба эти элементы поддерживаются соединительнотканными структурами, которые сохраняют протранственные взаимоотношения. Такие рецепторы называются вторичными (Рис.17). 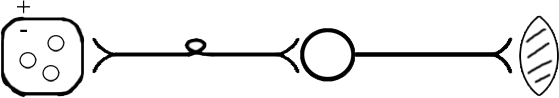 Рис.17. Структура вторичного рецептора Иногда функцию рецептора выполняет нервное окончание. Такой рецептор называется первичным (Рис.18). 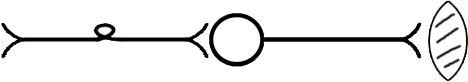 Рис.18. Структура первичного рецептора КЛАССИФИКАЦИЯ РЕЦЕПТОРОВ По месту расположения рецепторы бывают: Экстерорецепторы (расположены на коже и поверхностных слизистых). Интерорецепторы (расположены на внутренних органах). Проприорецепторы (в опорно-двигательном аппарате –мышцах,связках,сухожилиях). ЭКСТЕРОРЕЦЕПТОРЫ (контактные): Тактильные ( прикосновение, давление, вибрация). Терморецепторы (холодовые, тепловые). Вкусовые. Ноцерецепторы ( болевые). ЭКСТЕРОРЕЦЕПТОРЫ (дистантные). Обонятельные (хеморецепторы, реагирующие на запахи). Фоторецепторы (реакция на свет в сетчатке глаза). Фонорецепторы (реакция на звук в кортиевом органе). ИНТЕРОРЕЦЕПТОРЫ. Прессо или барорецепторы. Хеморецепторы. Терморецепторы. ноцерецепторы. СВОЙСТВА РЕЦЕПТОРОВ АДЕКВАТНОСТЬ – способность реагировать на специфический раздражитель, созданный для этого рецептора в эволюции (способность глаза реагировать на свет). Наиболее адекватны экстерорецепторы. ПОЛИМОДАЛЬНОСТЬ – способность рецептора реагировать на любые раздражители. Полимодальностью в большей степени обладают интерорецепторы. СПОСОБНОСТЬ отвечать серией импульсов в ответ на раздражение. От одних рецепторов идут очень частые импульсы, от других редкие, от третьих залпами. Благодаря этому нервная система может отличать раздражители (болевые, тактильные и т.д.). Чем сильнее раздражитель, тем более частые импульсы идут в нервный центр. СПОСОБНОСТЬ рецептора трансформировать энергию раздражителя в нервный импульс. Мембранный потенциал специализированной рецепторной клетки -55 мв.(ниже, чем для нерва и мышцы). Цитоплазма имеет заряд отрицательный, околоклеточная среда – положительный. На поверхности клетки преобладают ионы натрия. В цитоплазме находятся вакуоли с ацетилхолином (не у всех). При действии раздражителя частично увеличивается проницаемость мембраны для ионов натрия. Они частично поступают в цитоплазму рецепторной клетки и вызывают частичную деполяризацию, которая называется РЕЦЕПТОРНЫЙ ПОТЕНЦИАЛ (Рис.19). По характеру он является локальным током, может суммироваться и подчиняться закону «Силовых отношений», не достигает критического уровня деполяризации. 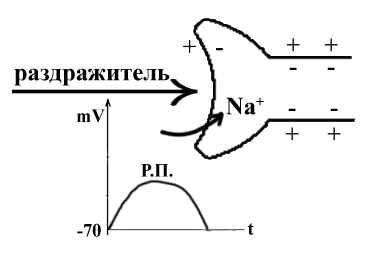 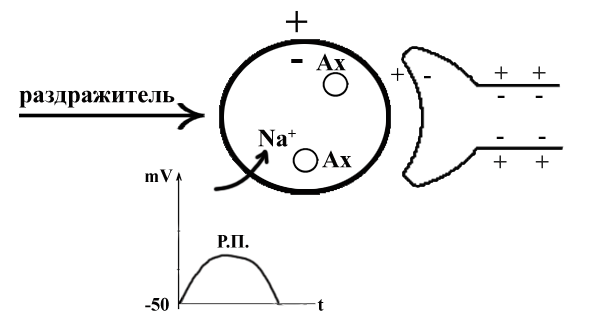 Рис. 19. Возникновение рецепторного потенциала в первичном и вторичном Рис. 19. Возникновение рецепторного потенциала в первичном и вторичномрецепторе. СПОНТАННАЯ АКТИВНОСТЬ – способность рецептора самовозбуждаться без действия раздражителя. Причиной является высокий тонус симпатических волокон вегетативной нервной системы. Эта система иннервирует все органы и клетки, регулируя уровень обменных процессов. Рецепторная клетка тоже получает подобную регуляцию. Из окончаний симпатических нервных волокон выделяется норадреналин, который увеличивает проницаемость мембраны рецептора к ионам натрия (Рис.20). Они уходят в цитоплазму и вызывают частичную деполяризацию (рецепторный потенциал) без действия раздражителя. При высоком рецепторном потенциале нервный центр будет воспринимать его как раздражитель. 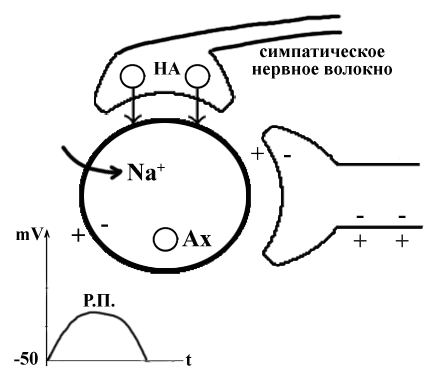 Рис.20.Механизм возникновения спонтанной активности рецептора. 6. ФЛЮКТУАЦИЯ – способность рецептора менять степень своей возбудимости. Зависит от тонуса волокон вегетативной нервной системы. При высоком тонусе симпатических нервных волокон из окончаний выделяется много норадреналина, возникает частичная деполяризация в рецепторе и его можно возбудить слабым раздражителем. Такая ситуация возникает днём, когда тонус симпатического отдела вегетативной системы высок, а рецепторы высоковозбудимы. При низком тонусе волокон симпатического отдела вегетативной нервной системы исходной частичной деполяризации в рецепторе не происходит, поэтому рецептор возбудить сложно и он является низковозбудимым. Например, ночью наступает царство «вагуса», т.е. повышается тонус парасимпатического отдела ВНС, рецепторы низковозбудимы и поэтому спящего человека трудно разбудить. Флюктуация позволяет рецепторам отдыхать. 7. АДАПТАЦИЯ – способность рецептора приспосабливаться к силе длительно действующего раздражителя. Наибольшей способностью к адаптации владеют экстерорецепторы и почти не обладают ею нтерорецепторы. Причины адаптации: А. При длительном действии раздражителя уменьшается проницаемость мембраны в рецепторе к ионам натрия. За счёт этого уменьшается высота рецепторного потенциала и частота импульсов, идущих по нервному волокну. Б. Возникает стойкая деполяризация в рецепторе, которая не меняется реполяризацией. В. Могут подключаться тормозные нейроны, которые выделяя ГАМК, вызывают гиперполяризацию. Г. При длительном действии раздражителя расходуется ацетилхолин, содержащийся во вторичном рецепторе. Все перечисленные причины адаптации не обязательны для каждого рецептора. |