Экзамен, который я обязательно сдам). Определение биологии как науки. Связь биологии с другими науками. Значение биологии для медицины. Фундаментальные свойства живого
 Скачать 1.73 Mb. Скачать 1.73 Mb.
|

|
Клетка – элементарная структурно-функциональная биологическая единица. Особенности строения эукариотической клетки. Клетка - элементарная единица живой системы. Элементарной единицей она может быть названа потому, что в природе нет более мелких систем, которым были бы присущи все без исключения признаки (свойства) живого. Известно, что организмы бывают одноклеточными (например, бактерии, простейшие, некоторые водоросли) или многоклеточными. Клетка обладает всеми свойствами живой системы: она осуществляет обмен веществ и энергии, растет, размножается и передает по наследству свои признаки, реагирует на внешние раздражители и способна двигаться. Она является низшей ступенью организации, обладающей всеми этими свойствами. Специфические функции в клетке распределены между органоидами, внутриклеточными структурами, имеющими определенную форму, такими, как клеточное ядро, митохондрии и др. У многоклеточных организмов разные клетки (например, нервные, мышечные, клетки крови у животных или клетки стебля, листьев, корня у растений) выполняют разные функции и поэтому различаются по структуре. Несмотря на многообразие форм, клетки разных типов обладают поразительным сходством в своих главных структурных особенностях. Все организмы, имеющие клеточное строение, делятся на две группы: предъядерные (прокариоты) и ядерные (эукариоты). Клетки прокариот, к которым относятся бактерии, в отличие от эукариот, имеют относительно простое строение. В прокариотической клетке нет организованного ядра, в ней содержится только одна хромосома, которая не отделена от остальной части клетки мембраной, а лежит непосредственно в цитоплазме. Однако в ней также записана вся наследственная информация бактериальной клетки. 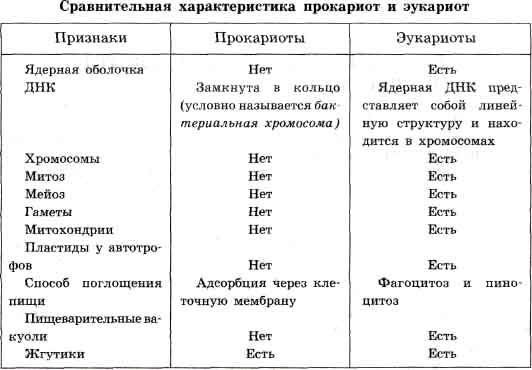 Для растительной клетки характерно наличие различных пластид, крупной центральной вакуоли, которая иногда отодвигает ядро к периферии, а также расположенной снаружи плазматической мембраны клеточной стенки, состоящей из целлюлозы. В клетках высших растений в клеточном центре отсутствует центриоль, встречающаяся только у водорослей. Резервным питательным углеводом в клетках растений является крахмал. В клетках представителей царства грибов клеточная стенка обычно состоит из хитина - вещества, из которого построен наружный скелет членистоногих животных. Имеется центральная вакуоль, отсутствуют пластиды. Только у некоторых грибов в клеточном центре встречается центриоль. Запасным углеводом в клетках грибов является гликоген. В клетках животных отсутствует плотная клеточная стенка, нет пластид. Нет в животной клетке и центральной вакуоли. Центриоль характерна для клеточного центра животных клеток. Резервным углеводом в клетках животных также является гликоген. Клеточная теория. История и современное состояние. Значение клеточной теории для биологии и медицины. Основные положения клеточной теории, ее значение Все живые организмы состоят из клеток — из одной клетки (одноклеточные организмы) или многих (многоклеточные). Клетка — это один из основных структурных, функциональных и воспроизводящих элементов живой материи; это элементарная живая система. Существуют неклеточные организмы (вирусы), но они могут размножаться только в клетках. Существуют организмы, вторично потерявшие клеточное строение (некоторые водоросли). История изучения клетки связана с именами ряда ученых. Р. Гук впервые применил микроскоп для исследования тканей и на срезе пробки и сердцевины бузины увидел ячейки, которые и назвал клетками. Антони ван Левенгук впервые увидел клетки под увеличением в 270 раз. М. Шлейден и Т. Шванн явились создателями клеточной теории. Они ошибочно считали, что клетки в организме возникают из первичного неклеточного вещества. Позднее Р. Вирхов сформулировал одно из важнейших положений клеточной теории: «Всякая клетка происходит из другой клетки...» Значение клеточной теории в развитии науки велико. Стало очевидно, что клетка — это важнейшая составляющая часть всех живых организмов. Она их главный компонент в морфологическом отношении; клетка является эмбриональной основой многоклеточного организма, т.к. развитие организма начинается с одной клетки — зиготы; клетка — основа физиологических и биохимических процессов в организме. Клеточная теория позволила прийти к выводу о сходстве химического состава всех клеток и еще раз подтвердила единство всего органического мира. Современная клеточная теория включает следующие положения: -клетка — основная единица строения и развития всех живых организмов, наименьшая единица живого; -клетки всех одноклеточных и многоклеточных организмов сходны (гомологичны) по своему строению, химическому составу, основным проявлениям жизнедеятельности и обмену веществ; -размножение клеток происходит путем их деления, и каждая новая клетка образуется в результате деления исходной (материнской) клетки; -в сложных многоклеточных организмах клетки специализированы по выполняемой ими функции и образуют ткани; из тканей состоят органы, которые тесно взаимосвязаны и подчинены нервным и гуморальным системам регуляции. 8. Гипотезы происхождения эукариотических клеток. Теория симбиогенеза. Наиболее популярна в настоящее время симбиотическая гипотеза происхождения эукариотических клеток, согласно которой основой, или клеткой-хозяином, в эволюции клетки эукариотического типа послужил анаэробный прокариот, способный лишь к амебоидному движению. Переход к аэробному дыханию связан с наличием в клетке митохондрии, которые произошли путем изменений симбионтов — аэробных бактерий, проникших в клетку-хозяина и сосуществовавших с ней. Сходное происхождение предполагают для жгутиков, предками которых служили симбионты-бактерии, имевшие жгутик и напоминавшие современных спирохет. Приобретение клеткой жгутиков имело наряду с освоением активного способа движения важное следствие общего порядка. Предполагают, что базальные тельца, которыми снабжены жгутики, могли эволюционировать в центриоли в процессе возникновения механизма митоза. Способность зеленых растений к фотосинтезу обусловлена присутствием в их клетках хлоропластов. Сторонники симбиотической гипотезы считают, что симбионтами клетки-хозяина, давшими начало хлоропластам, послужили прокариотические синезеленые водоросли. Серьезным доводом в пользу симбиотического происхождения митохондрий, центриолей и хлоропластов является то, что перечисленные органеллы имеют собственную ДНК. Вместе с тем белки бациллин и тубулин, из которых состоят жгутики и реснички соответственно современных прокариот и эукариот, имеют различное строение. Центральным и трудным для ответа является вопрос о происхождении ядра. Предполагают, что оно также могло образоваться из симбионта-прокариота. Увеличение количества ядерной ДНК, во много раз превышающее в современной эукариотической клетке ее количество в митохондрий или хлоропласте, происходило, по-видимому, постепенно путем перемещения групп генов из геномов симбионтов. Нельзя исключить, однако, что ядерный геном формировался путем наращивания генома клетки-хозяина (без участия симбионтов). Согласно инвагинационной гипотезе, предковой формой эукариотической клетки был аэробный прокариот. Внутри такой клетки-хозяина находилось одновременно несколько геномов, первоначально прикреплявшихся к клеточной оболочке. Органеллы, имеющие ДНК, а также ядро, возникли путем впячивания и отшнуровывания участков оболочки с последующей функциональной специализацией в ядро, митохондрий, хлоропласты. В процессе дальнейшей эволюции произошло усложнение ядерного генома, появилась система цитоплазматических мембран. Инвагинационная гипотеза хорошо объясняет наличие в оболочках ядра, митохондрий, хлоропластов, двух мембран. Однако она не может ответить на вопрос, почему биосинтез белка в хлоропластах и митохондриях в деталях соответствует таковому в современных прокариотических клетках, но отличается от биосинтеза белка в цитоплазме эукариотической клетки. Клонирование. В биологии - метод получения нескольких идентичных организмов путем бесполого (в том числе вегетативного) размножения. Именно так, на протяжении миллионов лет, размножаются в природе многие виды растений и некоторых животных. Однако сейчас термин "клонирование" обычно используется в более узком смысле и означает копирование клеток, генов, антител и даже многоклеточных организмов в лабораторных условиях. Появившиеся в результате бесполого размножения экземпляры по определению генетически одинаковы, однако и у них можно наблюдать наследственную изменчивость, обусловленную случайными мутациями или создаваемую искусственно лабораторными методами. Термин "клон" как таковой происходит от греческого слова "klon", что означает - веточка, побег, черенок, и имеет отношение, прежде всего, к вегетативному размножению. Клонирование растений черенками, почками или клубнями в сельском хозяйстве известно уже тысячи лет. При вегетативном размножении и при клонировании гены не распределяются по потомкам, как в случае полового размножения, а сохраняются в полном составе. Только у животных все происходит иначе. По мере роста клеток животных происходит их специализация, то есть клетки теряют способность реализовывать всю генетическую информацию, заложенную в ядре многих поколений. Клетка как открытая система. Организация потоков вещества, энергии в клетке. Специализация и интеграция клеток многоклеточного организма. Клетка — открытая система, поскольку ее существование возможно только в условиях постоянного обмена веществом и энергией с окружающей средой. Жизнедеятельность клетки обеспечивается процессами, образующими три потока: информации, энергии веществ. Благодаря наличию потока информации клетка приобретает структуру, отвечающую критериям живого, поддерживает ее во времени, передает в ряду поколений. В этом потоке участвуют ядро, макро молекулы, переносящие информацию в цитоплазму (мРНК), цитоплазматический аппарат транскрипции (рибосомы и полисомы, тРНК, ферменты активации аминокислот). Позже полипептиды, синтезированные на полисомах, приобретают третичную и четвертичную структуру, и используется в качестве катализаторов или структурных белков. Также функционируют геномы митохондрий, а в зеленых растениях — и хлоропластов. Поток энергии обеспечивается механизмами энергообеспечения — брожением, фото — или хемосинтезом, дыханием. Дыхательный обмен включает реакции расщепления низкокалорийного органического «топлива» в виде глюкозы, жирных кислот, аминокислот, использование выделяемой энергии для образования высококалорийного клеточного «топлива» в виде аденозинтрифосфата (АТФ). Энергия АТФ в разнообразных процессах преобразуется в тот или иной вид работы — химическую (синтезы), осмотическую (поддержание перепадов концентрации веществ), электрическую, механическую, регуляторную. Анаэробный гликолиз — процесс бескилородного расщепления глюкозы. Фотосинтез — механизм преобразования энергии солнечного света в энергию химических связей органических веществ. Клеточный цикл, его периодизация. Митотический цикл и его механизмы. Проблемы клеточной пролиферации в медицине. Повторяющаяся совокупность событий, обеспечивающих деление эукариотических клеток, получила название клеточного цикла. Продолжительность клеточного цикла зависит от типа делящихся клеток. Некоторые клетки, например, нейроны человека, после достижения стадии терминальной дифференцировки прекращают свое деление вообще. Клетки легких, почек или печени во взрослом организме начинают делиться лишь в ответ на повреждение соответствующих органов. Клетки эпителия кишечника делятся на протяжении всей жизни человека. Даже у быстро пролиферирующих клеток подготовка к делению занимает около 24 ч. Клеточный цикл разделяют на стадии : Митоз - М-фаза, деление клеточного ядра. G1 -фаза период перед синтезом ДНК. S-фаза - период синтеза (репликации ДНК). G2-фаза - период между синтезом ДНК и митозом. Интерфаза - период, включающий в себя G1 -, S- и G2-фазы. Цитокинез - деление цитоплазмы. Точка рестрикции, R-point - время в клеточном цикле, когда продвижение клетки к делению становится необратимым. G0 фаза - состояние клеток, достигших монослоя или лишенных фактора роста в ранней G1 фазе.Делению клетки (митозу или мейозу) предшествует удвоение хромосом, которое происходит в периоде S клеточного цикла. Период обозначают первой буквой слова synthesis - синтез ДНК. С момента окончания периода S до завершения метафазы ядро содержит в четыре раза больше ДНК, чем ядро сперматозоида или яйцеклетки, а каждая хромосома состоит из двух идентичных сестринских хроматид. Во время митоза хромосомы конденсируются и в конце профазы или начале метафазы становятся различимыми при оптической микроскопии. Для цитогенетического анализа обычно используют препараты именно метафазных хромосом. В начале анафазы центромеры гомологичных хромосом разъединяются, и хроматиды расходятся к противоположным полюсам митотического веретена. После того как к полюсам отойдут полные наборы хроматид (с этого момента их называют хромосомами), вокруг каждого из них образуется ядерная оболочка, формируя ядра двух дочерних клеток (разрушение ядерной оболочки материнской клетки произошло в конце профазы ). Дочерние клетки вступают в период G1 , и только при подготовке к следующему делению они переходят в период S и в них происходит репликация ДНК. Клетки со специализированными функциями, длительное время не вступающие в митоз или вообще утратившие способность к делению, находятся в состоянии, называемом периодом G0 . Большинство клеток в организме диплоидные - то есть имеют два гаплоидных набора хромосом (гаплоидный набор - это число хромосом в гаметах, у человека он составляет 23 хромосомы, а диплоидный набор хромосом - 46). В гонадах предшественники половых клеток сначала претерпевают ряд митотических делений, а затем вступают в мейоз - процесс образования гамет, состоящий из двух последовательных делений. В мейозе гомологичные хромосомы спариваются (отцовская 1-я хромосома с материнской 1-й хромосомой и т. д.), после чего в ходе так называемого кроссинговера происходит рекомбинация, то есть обмен участками между отцовской и материнской хромосомами. В результате качественно изменяется генетический состав каждой из хромосом. В первом делении мейоза расходятся гомологичные хромосомы (а не сестринские хроматиды, как в митозе), вследствие чего образуются клетки с гаплоидным набором хромосом, каждая из которых содержит по 22 удвоенные аутосомы и одной удвоенной половой хромосоме. Между первым и вторым делениями мейоза нет периода S, а в дочерние клетки во втором делении расходятся сестринские хроматиды. В итоге образуются клетки с гаплоидным набором хромосом, в которых вдвое меньше ДНК, чем в диплоидных соматических клетках в периоде G1, и в 4 раза меньше - чем в соматических клетках по окончании периода S. При оплодотворении число хромосом и содержание ДНК у зиготы становится таким же, как в соматической клетке в периоде G1. Период S в зиготе открывает путь к регулярному делению, характерному для соматических клеток. Митоз (от греч. mitos - нить) - деление ядра, следующее за репликацией хромосом, в результате чего дочерние ядра содержат то же число хромосом, что и родительские. Митоз имеет сложный механизм, включающий несколько фаз, необходимость которого возникла в процессе эволюции тогда, когда появились клетки с резко увеличенным количеством ДНК, упакованной в отдельные хромосомы. Процесс митоза составляют: профаза, прометафаза, метафаза, анафаза и телофаза. Профаза. В начале профазы многочисленные цитоплазматические микротрубочки, входящие в состав цитоскелета, распадаются; при этом образуется большой пул свободных молекул тубулина. Эти молекулы вновь используются для построения главного компонента митотического аппарата - митотического веретена. Каждая пара центриолей становится частью митотического центра, от которого лучами расходятся микротрубочки (фигура "звезда"). Вначале обе звезды лежат рядом около ядерной мембраны. В поздней профазе пучки полюсных микротрубочек, взаимодействующие друг с другом (и видимые в световой микроскоп как полюсные нити), удлиняются и как будто расталкивают два митотических центра друг от друга вдоль наружной поверхности ядра. Таким способом образуется биполярное митотическое веретено. Вторая стадия митоза - прометафаза начинается с быстрого распада ядерной оболочки на мелкие фрагменты, неотличимые от фрагментов цитоплазматического ретикулума. Эти фрагменты остаются видимыми около веретена. В клетках млекопитающих прометафаза занимает 10-20 минут. Расположенное около ядра митотическое веретено может теперь проникнуть в ядерную область. В хромосомах с каждой стороны центромеры образуются особые структуры - кинетохоры. Обычно у каждой хромосомы оказывается по одной кинетохорной нити, связанной с каждым из полюсов. В результате этого возникают две противоположно направленные силы, которые и приводят хромосому в экваториальную плоскость. Таким образом, беспорядочные прометафазные движения хромосом и их случайная окончательная ориентация обеспечивает случайную сегрегацию хроматид между дочерними клетками, столь важную в мейозе. Третья стадия митоза - метафаза часто продолжается длительное время. Все хромосомы располагаются таким образом, что их центромеры лежат в одной плоскости (метафазная пластинка). Метафазные хромосомы удерживаются в обманчиво статичном состоянии сбалансированными полярными силами. За ориентацию хромосом перпендикулярно оси митотического веретена и расположение их на равном расстоянии от обеих полюсов веретена, скорее всего, ответственны кинетохорные нити. Вероятно, такое расположение хромосом в метофазной пластинке обусловлено способом создания тянущей силы в митотическом веретене: этот способ таков, что сила, действующая на кинетохорные нити тем слабее, чем ближе к полюсу находятся кинетохоры . см. метафаза 1 и 2. Каждая хромосома удерживается в метафазной пластинке парой кинетохоров и двумя пучками связанных с ними нитей, идущих к противоположным полюсам веретена. Метафаза резко оканчивается разделением двух кинетохоров каждой хромосомы. Четвертая стадия митоза - анафаза продолжается обычно всего несколько минут. Анафаза начинается внезапным расщеплением каждой хромосомы, которое обусловлено разделением сестринских хроматид в точке их соединения в центромере. Это расщепление, разделяющее кинетохоры , не зависит от других событий митоза и происходит даже в хромосомах, не прикрепленных к митотическому веретену; оно позволяет полярным силам веретена, действующим на метафазную пластинку, начать перемещение каждой хроматиды к соответствующим полюсам веретена со скоростью порядка 1 мкм/мин. Во время этого анафазного движения кинетохорные нити укорачиваются по мере того, как хромосомы приближаются к полюсам. Примерно в это же время удлиняются нити митотического веретена и два полюса веретена расходятся еще дальше. Далее см. Митоз: движение хромосом в анафазе Клеточная стадия, на которой хромосомы расходятся к двум полюсам новых дочерних клеток. В пятой заключительной стадии митоза телофазе разделенные дочерние хроматиды подходят к полюсам, кинетохорные нити исчезают. После удлинения полюсных нитей вокруг каждой группы дочерних хроматид образуется новая ядерная оболочка. Конденсированный хроматин начинает разрыхляться, появляются ядрышки, и митоз заканчивается. Пролиферация. Основной способ деления тканевых клеток — это митоз. По мере увеличения числа клеток возникают клеточные группы, или популяции, объединенные общностью локализации в составе зародышевых листков (эмбриональных зачатков) и обладающие сходными гистогенетическими потенциями. Клеточный цикл регулируется многочисленными вне- и внутриклеточными механизмами. К внеклеточным относятся влияния на клетку цитокинов, факторов роста, гормональных и нейрогенных стимулов. Роль внутриклеточных регуляторов играют специфические белки цитоплазмы. В течение каждого клеточного цикла существуют несколько критических точек, соответствующих переходу клетки из одного периода цикла в другой. При нарушении внутренней системы контроля клетка под влиянием собственных факторов регуляции элиминируется апоптозом, либо на некоторое время задерживается в одном из периодов цикла. |