Экзамен, который я обязательно сдам). Определение биологии как науки. Связь биологии с другими науками. Значение биологии для медицины. Фундаментальные свойства живого
 Скачать 1.73 Mb. Скачать 1.73 Mb.
|

|
Тип аддитивного действия генов, каждый из которых оказывает свою, часто небольшую, долю влияния на признак, называется кумулятивной полимерией. Используя решетку Пеннета, можно подсчитать частоты доминантных генов среди генотипов второго поколения. Для этого в каждой из 64 клеток вместо генотипа записывается число присутствующих в нем доминантных аллелей. Определив частоты доминантных аллелей, можно убедиться, что генотипы с числом доминантных генов 6,5,4,3, 2, 1,0 встречаются 1,6,15,20,15,6,1 раз соответственно. 38.Наследование групп крови и резус-фактора. Группа крови На эритроцитах имеются специальные белки - антигены групп крови. В плазме к этим антигенам имеются антитела. При встрече одноименных антигена и антитела происходит их взаимодействие и склеивание эритроцитов в монетные столбики. В таком виде они не могут переносить кислород. Поэтому в крови одного человека не встречаются одноименные антиген и антитело. Их комбинация - группа крови.Антигены и антитела групп крови, как все белки организма, наследуются - именно белки, а не сами группы крови, поэтому комбинация этих белков у детей может отличаться от комбинации у родителей и получаться другая группа крови. Существует множество антигенов на эритроцитах и множество систем групп крови. В рутинной диагностике пользуются определением группы крови по системе АВ0. Антигены: А, В; антитела: альфа, бета. Наследование: ген IA кодирует синтез белка А, IB - белка В, i не кодирует синтез белков. Группа крови I (0). Генотип ii. Отсутствие антигенов на эритроцитах, присутствие обоих антител в плазме Группа крови II (А). Генотип IA\IA или IА\i. Антиген А на эритроцитах, антитело бета в плазме Группа крови III (В). Генотип IB\IB или IВ\i. Антиген В на эритроцитах, антитело альфа в плазме Группа крови IV (АВ). Генотип IA\IB. Оба антигена на эритроцитах, отсутствие антител в плазме. Наследование: У родителей с первой группой крови может родиться ребенок только с первой группой. У родителей со второй - ребенок с первой или второй. У родителей с третьей - ребенок с первой или третьей. У родителей с первой и второй - ребенок с первой или второй. У родителей с первой и третьей - ребенок с первой или третьей. У родителей с второй и третьей - ребенок с любой группой крови. У родителей с первой и четвертой - ребенок с второй и третьей. У родителей с второй и четвертой - ребенок с второй, третьей и четвертой У родителей с третьей и четвертой - ребенок с второй, третьей и четвертой. У родителей с четвертой - ребенок с второй, третьей и четвертой. Если у одного из родителей первая группа крови, у ребенка не может быть четвертой. И наоборот - если у одного из родителей четвертая, у ребенка не может быть первой. Групповая несовместимость: При беременности может возникнуть не только резус-конфликт, но и конфликт по группам крови. Если плод имеет антиген, которого нет у матери, она может вырабатывать против него антитела: антиА, антиВ. Конфликт может возникнуть если плод имеет II группу крови, а мать I или III; плод III, а мать I или II; плод IV, а мать любую другую. Нужно проверять наличие групповых антител во всех парах, где у мужчины и женщины разные группы крови, за исключением случаев, когда у мужчины первая группа. Резус-фактор Белок на мембране эритроцитов. Присутствует у 85% людей - резус-положительных. Остальные 15% - резус-отрицательны. Наследование: R- ген резус-фактора. r - отсутствие резус фактора. Родители резус-положительны (RR, Rr) - ребенок может быть резус-положительным (RR, Rr) или резус-отрицательным (rr). Один родитель резус-положительный (RR, Rr), другой резус-отрицательный (rr) - ребенок может быть резус-положительным (Rr) или резус-отрицательным (rr). Родители резус-отрицательны, ребенок может быть только резус-отрицательным. Резус-фактор, как и группу крови, необходимо учитывать при переливании крови. При попадании резус фактора в кровь резус-отрицательного человека, к нему образуются антирезусные антитела, которые склеивают резус-положительные эритроциты в монетные столбики. Резус-конфликт Может возникнуть при беременности резус-отрицательной женщины резус-положительным плодом (резус-фактор от отца). При попадании эритроцитов плода в кровоток матери, против резус-фактора у нее образуются антирезусные антитела. В норме кровоток матери и плода смешивается только во время родов, поэтому теоретически возможным резус-конфликт считается во вторую и последующие беременности резус-положительным плодом. Практически в современных условиях часто происходит повышение проницаемости сосудов плаценты, различные патологии беременности, приводящие к попаданию эритроцитов плода в кровь матери и во время первой беременности. Поэтому антирезусные антитела необходимо определять при любой беременности у резус-отрицательной женщины начиная с 8 недель (время образования резус-фактора у плода). Для предотвращения их образования во время родов, в течение 72 часов после любого окончания беременности срока более 8 недель вводят антирезусный иммуноглобулин. 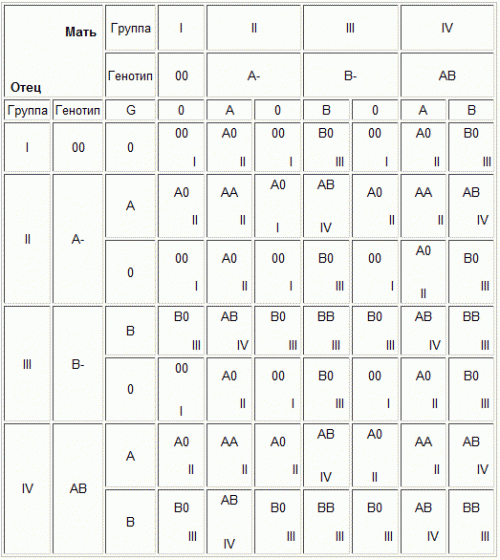 40. Взаимодействие неаллельных генов. Комплементарность, эпистаз, полимерия. Явление плейотропии. Неаллельные гены- это гены, расположенные в различных участках хромосом и кодирующие неодинаковые белки. Неаллельные гены также могут взаимодействовать между собой. При этом либо один ген обусловливает развитие нескольких признаков, либо, наоборот, один признак проявляется под действием совокупности нескольких генов. Выделяют три формы и взаимодействия неаллельных генов: комплементарность; полимерия; эпистаз; плейотропия. Комплемента́рное (дополнительное) действие генов — это вид взаимодействия неаллельных генов, доминантные аллели которых при совместном сочетании в генотипе обусловливают новое фенотипическое проявление признаков. Полимери́я — взаимодействие неаллельных множественных генов, однозначно влияющих на развитие одного и того же признака; степень проявления признака зависит от количества генов. Полимерные гены обозначаются одинаковыми буквами, а аллели одного локуса имеют одинаковый нижний индекс. Эписта́з — взаимодействие неаллельных генов, при котором один из них подавляется другим. Подавляющий ген называется эпистатичным, подавляемый — гипостатичным. Если эпистатичный ген не имеет собственного фенотипического проявления, то он называется ингибитором и обозначается буквой I. Эпистатическое взаимодействие неаллельных генов может быть доминантным и рецессивным. При доминантном эпистазе проявление гипостатичного гена (В, b) подавляется доминантным эпистатичным геном (I > В, b). Расщепление по фенотипу при доминантном эпистазе может происходить в соотношении 12:3:1, 13:3, 7:6:3. Рецессивный эпистаз — это подавление рецессивным аллелем эпистатичного гена аллелей гипостатичного гена (i > В, b). Расщепление по фенотипу может идти в соотношении 9:3:4, 9:7, 13:3. 53 Плейотропное действие гена. Первичная и вторичная плейотропия. Летальные гены. Примеры. Плейотропное действие генов - это зависимость нескольких признаков от одного гена, то есть множественное действие одного гена. Выражается в способности одного гена влиять на несколько фенотипических признаков. Таким образом, новая мутация в гене может оказать влияние на некоторые или все связанные с этим геном признаки. Ген, обуславливающий рыжие волосы, обуславливает более светлую окраску кожи и появление веснушек. Фенилкетонурия (ФКУ), болезнь, вызывающая задержку умственного развития, выпадение волос и пигментацию кожи, может быть вызвана мутацией в гене, кодирующем ферментфенилаланин-4-гидроксилаза, который в норме катализирует превращение аминокислоты фенилаланина в тирозин. Арахнодактилия, вызываемая доминантной мутацией, проявляется одновременно в изменениях пальцев рук и ног, вывихах хрусталика глаза и врождённых пороках сердца. При первичной плейотропии ген проявляет свой множественный эффект. Например, при болезни Хартнупа мутация гена приводит к нарушению всасывания аминокислоты триптофана в кишечнике и его реабсорбции в почечных канальцах. При этом поражаются одновременно мембраны эпителиальных клеток кишечника и почечных канальцев с расстройствами пищеварительной и выделительной систем. При вторичной плейотропии есть один первичный фенотипический проявление гена, вслед за которым развивается ступенчатый процесс вторичных изменений, приводящих к множественным эффектам. Так, при серповидно клеточной анемии у гомозигот наблюдается несколько патологических признаков: анемия, увеличенная селезенка, поражение кожи, сердца, почек и мозга. Поэтому гомозиготы с геном серповидно клеточной анемии гибнут, как правило, в детском возрасте. Все эти фенотипные проявления гена составляют иерархию вторичных проявлений +Летальными генами называются гены, носители которых погибают из-за нарушений развития или заболеваний, связанных с работой данного гена, фенотипический эффект которых вызывает гибель организма при определенных условиях, или на определенных этапах развития чаще всего на эмбриональныхстадиях развития. 41. Механизмы генотипического определения и дифференциации признака пола в развитии. Половой диморфизм. Начало изучению генотипического определения пола было положено открытием американскими цитологами у насекомых различия в форме, а иногда и в числе хромосом у особей разного пола (Мак-Кланг, 1906, Уилсон, 1906) и классическими опытами немецкого генетика Корренса по скрещиванию однодомного и двудомного видов брионии. Уилсон обнаружил, что у клопа Lydaeus turucus самки имеют 7 пар хромосом, у самцов же 6 пар одинаковых с самкой хромосом, а в седьмой паре одна хромосома такая же, как соответствующая хромосома самки, а другая маленькая. Пара хромосом, которые у самца и самки разные, получила название идио, или гетерохромосомы, или половые хромосомы. У самки две одинаковые половые хромосомы, обозначаемые как Х-хромосомы, у самца одна Х-хромосома, другая - Y-хромосома. Остальные хромосомы одинаковые у самца и у самки, были названы аутосомами. Таким образом, хромосомная формула у самки названного клопа запишется 12A + XX, у самца 2A + XY. У ряда других организмов, хотя и существует в принципе тот же аппарат для определения пола, однако гетерозиготны в отношении реализаторов пола не мужские, а женские организмы. Особи мужского пола имеют две одинаковые половые хромосомы ZZ, а особи женского пола - ZO или ZW. ZZ-ZW тип определения пола наблюдается у бабочек, птиц, ZZ-ZO - ящериц, некоторых птиц. Совершенно другой механизм определения пола, называемый гаплодиплоидный, широко распространен у пчел и муравьев. У этих организмов нет половых хромосом: самки - это диплоидные особи, а самцы (трутни) - гаплоидные. Самки развиваются из оплодотворенныз яиц, а из неоплодотворенных развиваются трутни. Человек в отношении определения пола относится к типу XX-XY. При гаметогенезе наблюдается типичное менделевское расщепление по половым хромосомам. каждая яйцеклетка содержит одну Х-хромосому, а другая половина - одну Y-хромосому. Пол потомка зависит от того, какой спермий оплодотворит яйцеклетку. Пол с генотипом ХХ называют гомогаметным, так как у него образуются одинаковые гаметы, содержащие только Х-хромосомы, а пол с генотипом XY-гетерогаметным, так как половина гамет содержит Х-, а половина - Y-хромосому. У человека генотипический пол данного индивидума определяют, изучая неделящиеся клетки. Одна Х-хромосома всегда оказывается в активном состоянии и имеет обычный вид. Другая, если она имеется, бывает в покоящемся состоянии в виде плотного темно-окрашенного тельца, называемого тельцем Барра (факультативный гетерохроматин). Число телец Барра всегда на единицу меньше числа наличных х-хромосом, т.е. в мужском организме их нет вовсе, у женщин (ХХ) - одно. У человека Y-хромосома является генетически инертной, так как в ней очень мало генов. Однако влияние Y-хромосомы на детерминацию пола у человека очень сильное. Хромосомная структура мужчины 44A+XY и женщины 44A+XX такая же, как и у дрозофилы, однако у человека особь кариотипом 44A+XD оказалась женщиной, а особь 44A+XXY мужчиной. В обоих случаях они проявляли дефекты развития, но все же пол определялся наличием или отсутствием y-хромосомы. Люди генотипа XXX2A представляют собой бесплодную женщину, с генотипом XXXY2A - бесплодных умственно отстающих мужчин. Такие генотипы возникают в результате нерасхождения половых хромосом, что приводит к нарушению развития (например, синдром Клайнфельтера (XXY). Нерасхождение хромосом изучаются как в мейозе, так и в митозе. Нерасхождение может быть следствием физического сцепления Х-хромосом, в таком случае нерасхождение имеет место в 100% случаев. Всем млекопитающим мужского пола, включая человека, свойственен так называемый H-Y антиген, находящийся на поверхности клеток, несущих Y-хромосому. Единственной функцией его считается дифференцировка гонад. Вторичные половые признаки развиваются под влиянием стероидных гормонов, вырабатываемых гонадами. Развитие мужских вторичных половых признаков контролирует тестостерон, воздействующий на все клетки организма, включая клетки гонад. Мутация всего одного Х-хромосомы, кодирующего белок-рецептор тестостерона, приводит к синдрому тестикумерной фелинизации особей XY. Клетки-мутанты не чувствительны к действию тестостерона, в результате чего взрослый организм приобретает черты, характерные для женского пола. При этом внутренние половые органы оказываются недоразвитыми и такие особи полностью стерильные. Таким образом, в определении и дифференцировке пола млекопитающих и человека взаимодействуют хромосомный и генный механизмы. +Несмотря на то, что женщины имеют две Х-хромосомы, а мужчины - только одну, экспрессия генов Х-хромосомы происходит на одном и том же уровне у обоих полов. Это объясняется тем, что у женщин в каждой клетке полностью инактивирована одна Х-хромосома (тельце Барра), о чем уже было сказано выше. Х-хромосома инактивируется на ранней стадии эмбрионального развития, соответствующей времени имплантации. при этом в разных клетках отцовская и материнская Х-хромосомы выключаются случайно. Состояние инактивации данной Х-хромосомы наследуется в ряду клеточных делений. Таким образом, женские особи, гетерозиготные по генам половых хромосом, представляют собой мозаики (пример, черепаховые кошки).Таким образом, пол человека представляет собой менделирующий признак, наследуемый по принципу обратного (анализирующего) скрещивания. Гетерозиготой оказывается гетерогаметный пол (XY), который скрещивается с рецессивной гомозиготой, представленной гомогаметным полом (XX). 42. Наследование признаков человека, сцепленных с полом. Морган и его сотрудники заметили, что наследование окраски глаз у дрозофилы зависит от пола родительских особей, несущих альтернативные аллели. Красная окраска глаз доминирует над белой. При скрещивании красноглазого самца с белоглазой самкой в F1, получали равное число красноглазых самок и белоглазых самцов. Однако при скрещивании белоглазого самца с красноглазой самкой в F1 были получены в равном числе красноглазые самцы и самки При скрещивании этих мух F1, между собой были получены красноглазые самки, красноглазые и белоглазые самцы, но не было ни одной белоглазой самки. Тот факт, что у самцов частота проявления рецессивного признака была выше, чем у самок, наводил на мысль, что рецессивный аллель, определяющий белоглазость, находится в Х - хромосоме, а Y - хромосома лишена гена окраски глаз. Чтобы проверить эту гипотезу, Морган скрестил исходного белоглазого самца с красноглазой самкой из F1. В потомстве были получены красноглазые и белоглазые самцы и самки. Из этого Морган справедливо заключил, что только Х - хромосома несет ген окраски глаз. В Y - хромосоме соответствующего локуса вообще нет. Это явление известно под названием наследования, сцепленного с полом. Гены, находящиеся в половых хромосомах, называют сцепленными с полом. В Х-хромосоме имеется участок, для которого в Y-хромосоме нет гомолога. Поэтому у особей мужского пола признаки, определяемые генами этого участка, проявляются даже в том случае, если они рецессивны. Эта особая форма сцепления позволяет объяснить наследование признаков, сцепленных с полом. При локализации признаков как в аутосоме, так и в Х- b Y-хромосоме наблюдается полное сцепление с полом. У человека около 60 генов наследуются в связи с Х-хромосомой, в том числе гемофелия, дальтонизм (цветовая слепота), мускульная дистрофия, потемнение эмали зубов, одна из форм агаммглобулинемии и другие. Наследование таких признаков отклоняется от закономерностей, установленных Г.Менделем. Х-хромосома закономерно переходит от одного пола к другому, при этом дочь наследует Х-хромосому отца, а сын Х-хромосому матери. Наследование, при котором сыновья наследуют признак матери, а дочери - признак отца получило, название крисс-кросс (или крест-накрест). Известны нарушения цветового зрения, так называемая цветовая слепота. В основе появления этих дефектов зрения лежит действие ряда генов. Красно-зеленая слепота обычно называется дальтонизмом. Еще задолго до появления генетики в конце XVIII и в XIX в. было установлено, что цветовая слепота наследуется согласно вполне закономерным правилам. Так, если женщина, страдающая цветовой слепотой, выходит замуж за мужчину с нормальным зрением, то у их детей наблюдается очень своеобразная картина перекрестного наследования. Все дочери от такого брака получат признак отца, т.е. они имеют нормальное зрение, а все сыновья, получая признак матери, страдают цветовой слепотой (а-дальтонизм, сцепленный с Х-хромосомой) Р Ха Ха х Ха y Ха Ха,y F1 Ха Ха, Хаy В том же случае, когда наоборот, отец является дальтоником, а мать имеет нормальное зрение, все дети оказываются нормальными. В отдельных браках, где мать и отец обладают нормальным зрением, половина сыновей может оказаться пораженными цветовой слепотой. В основном наличие цветовой слепоты чаще встречается у мужчин. Э.Вильсон объяснил наследование этого признака, предположив, что он локализовал в Х-хромосоме и что у человека гетерогаметным (XY) является мужской пол. Становится вполне понятным, что в браке гомозиготной нормальной женщины (Ха Ха) с мужчиной дальтоником (Х аy) все дети рождаются нормальными. Однако при этом, все дочери становятся скрытыми носителями дальтонизма, что может проявиться в последующих поколениях. Другим примером наследования сцепленного с полом, может послужить рецессивныйполулетальный ген, вызывающий несвертываемость крови на воздухе - гемофилию. Это заболевание появляется почти исключительно только у мальчиков. При гемофилии нарушается образование фактора VIII, ускоряющего свертывание крови. ген, детерминирующий синтех фактора VIII, находится в участке Х- хромосомы, недоминантным нормальным и рецессивным мутантным. Возможны следующие генотипы и фенотипы: Генотипы Фенотипы Хн Хн Нормальная женщина Хн Хn Нормальная женщина (носитель) Хнy Нормальный мужчина Хny Мужчина гемофилик Один из сцепленных с полом рецессивных генов вызывает особый тип мышечной дистрофии (тип Дюмена). Эта дистрофия проявляется в раннем детстве и постепенно ведет к инвалидности и смерти ранее 20-летнего возраста. Потому мужчины с дистрофией Дюмена не имеют потомства, а женщины гетерозиготные по гену этого заболевания, вполне нормальны. Среди доминантных признаков, связанных с Х-хромосомой, можно указать на ген, который вызывает недостаточность органического фосфора в крови. В результате, при наличии этого гена, часто развивается рахит, устойчивый к лечению обычными дозами витамина А. В этом случае картина сцепленного с полом наследования заметно отличается от того хода передачи по поколениям, который был описан для рецессивных болезней. В браках девяти больных женщин со здоровыми мужчинами среди детей была половина больных девочек и половина мальчиков. Здесь, в соответствии с характером наследование доминантного гена, в Х-хромосомах произошло расщепление в отношении 1:1:1:1. Другим примером доминантного гена, локализованного в Х-хромосоме человека, может послужить ген, вызывающий дефект зубов, приводящий к потемнению эмали зубов. Так как гетерогаметный пол гемизиготен по сцепленным с полом генам, то эти гены всегда проявляются в их фенотипе, даже если они рецессивны. Большинство генов, имеющихся в Х-хромосоме, в Y-хромосоме отсутствует, однако определенную генетическую информацию она все-таки несет. Различают два типа такой информации: во-первых, содержащуюся в генах, присутствующих только в Y- хромосоме, и, во-вторых, в генах, присутствующих как в Y-, так и в Х- хромосоме (гемфрагический диатез). Y-хромосома передается от отца всем его сыновьям, и только им. Следовательно, для генов, содержащихся только в Y-хромосоме, характерно голандрическое наследование, т.е они передаются от отца к сыну и проявляются у мужского пола. У человека в Y-хромосоме содержатся по крайней мере три гена, один из которых необходим для дифференциации семенников, второй требуется для проявления антигена гистосовместимости, а третий оказывает влияние на размер зубов. Y-хромосома имеет немного признаков, среди которых есть патологические. Патологические признаки наследуются по параллельной схеме наследования (100%-ое проявление по мужской линии). К ним относят: 1) облысение; 2) гипертрихоз (оволосенение козелка ушной раковины в зрелом возрасте); 3) наличие перепонок на нижних конечностях; +4) ихтиоз (чешуйчатость и пятнистое утолщение кожи). |