Экзамен, который я обязательно сдам). Определение биологии как науки. Связь биологии с другими науками. Значение биологии для медицины. Фундаментальные свойства живого
 Скачать 1.73 Mb. Скачать 1.73 Mb.
|

|
28. Предмет, задачи, методы генетики. Этапы развития генетики. Вклад отечественных ученых в развитие генетики. Значение генетики для медицины. Генетика человека – основа медицинской генетики. Человек – удобный генетический объект. У человека лучше, чем у других видов изучены биохимические,иммунологические, физиологические и другие реакции, а эти признаки детерминированы генами. 98-99% заболеваний возникают в результате поражения генетического материала. 1-2% - травмы и ожоги. Генетика человека – наука о наследственно обусловленных различиях людей и о нарушениях генетического материала. ГЕНЕТИКИ. ЕВГЕНИКА История Развитие генетики происходило под влиянием полит. Сил. Платон в своем труде настаивал на том что потомки лучших представителей обоих полов должны воспит. Тщательно. По мнению демократа способности большинства людей развиваются за счет упражнения. Генетика чел-ка появ. в 1865 г. Законы менделя были использованы к человеку. С появлением молекулярной биологии использование методов биометрии пошло на убыль. 50-60 гг характеризуются активным исследование хромосом человека. В конце 60-70 гг появление методов пренатальной диагностики хромосомных болезней. Стало очевидно что легче предупредить чем лечить. Разделы генетики Биохимическая - изучает биохимию нуклеиновых кислот белков ферментов Цитогенетика - изучение хромосом в норме и при патологии Иммуногенетика - генетика групп крови Формальная - изучает наследование менделеевских признаков Клиническая - решает задачи диагностики, лечения наел болезней Популяционная-поведения генов в популяции генетика сомат клеток- перенос генов на клет уровне Методы генетики Совокупность методов исследования наследственных свойств организма (его генотипа) называется генетический анализ. В зависимости от задачи и особенностей изучаемого объекта генетический анализ проводят на популяционном, организменном, клеточном и молекулярном уровнях. Основу генетического анализа составляет гибридологический анализ, основанный на анализе наследования признаков при скрещиваниях. Гибридологический анализ, основы которого разработал основатель современной генетики Г. Мендель, основан на следующих принципах: 1. Использование в качестве исходных особей (родителей), форм, не дающих расщепления при скрещивании, т. е. константных форм. 2. Анализ наследования отдельных пар альтернативных признаков, то есть признаков, представленных двумя взаимоисключающими вариантами. 3. Количественный учет форм, выщепляющихся в ходе последовательных скрещиваний и использование математических методов при обработке результатов. 4. Индивидуальный анализ потомства от каждой родительской особи. 5. На основании результатов скрещивания составляется и анализируется схема скрещиваний. Этапы развития генетики этап классической генетики (Создана теория гена и хромосомной теории - наследственности) разработано учение о фенотипе и генотипе. этап неоклассицизма. Возможность экспериментального мутагенеза, ген дискретен, создана биохим генетика, ДНК - основа записи, развитие радиац генетики 1953-по сей день Эпоха синтетической генетики. Теория гена, теория кутации, в мед генетики имеется понятие кариотип, идиограмма, кариограмма. Евгеника (от греч. eugenes — хорошего рода), учение о наследственном здоровье человека и путях улучшения его наследственных свойств, о возможных методах активного влияния на эволюцию человечества в целях дальнейшего совершенствования его природы, об условиях и законах наследования одарённости й таланта, о возможном ограничении передачи наследственных болезней будущим поколениям. Основы современной генетики заложены Г. Менделем, открывшим законы дискретной наследственности (1865), и школой Т. Х. Моргана, обосновавшей хромосомную теорию наследственности (1910-е гг.). В СССР в х годах выдающийся вклад в генетику внесли работы, Н. К. Кольцова, С. С. Четверикова, А. С. Серебровского и др. Н. К. Кольцов выдвинул в 1927 концепцию о том, что хромосома с генами представляет одну гигантскую органическую молекулу и что воспроизведение этой наследственной молекулы осуществляется матричным путем. То и другое было позже подтверждено, когда генетические процессы начали изучать на молекулярном уровне (правда оказалось, что генетическим материалом служит не белок, как считал Кольцов, а ДНК). Серебровский является одним из основателей генетики популяций. Его учение о генофонде и геногеографии заложило оригинальное эволюционно-географическое направление в генетике и селекции. Концепция генофонда, глубоко проникшая в генетику, легла в основу природоохранной политики. Полученные им радиационные мутации послужили толчком для создания гипотезы появления структурных мутаций. Серебровский внес большой вклад в разработку основ селекции и гибридизации, методов генетического анализа. 29. Уровни организации наследственного материала. В наследственной структуре клетки и организма в целом выделяют три уровня организации генетического материала: генный, хромосомный и геномный. Генный уровень Наименьшей (элементарной) единицей наследственного материала является ген. Ген – это часть молекулы ДНК, имеющая определенную последовательность нуклеотидов и представляющая собой единицу функционирования наследственного материала. Ген несет информацию о конкретном признаке или свойстве организма. У человека имеется около 30 тысяч генов. Изменение в структуре гена ведет к изменению соответствующего признака. Следовательно, на генном уровне обеспечиваются индивидуальное наследование и индивидуальная изменчивость признаков. Хромосомный уровень Все гены в клетке объединены в группы и располагаются в хромосомах в линейном порядке. Каждая хромосома уникальна по набору входящих в нее генов. В состав хромосом входят ДНК, белки (гистоновые и негистоновые), РНК, полисахариды, липиды и ионы металлов. Хромосомный уровень в эукариотических клетках обеспечивает характер функционирования отдельных генов, тип их наследования и регуляцию их активности. Он позволяет закономерно воспроизводить и передавать наследственную информацию в процессе деления клетки. Геномный уровень Геном – совокупность всех генов, находящихся в гаплоидном наборе хромосом. При оплодотворении два генома родительских гамет сливаются и образуют генотип. Генотип – совокупность всех генов, заключенных в диплоидном наборе хромосом, или кариотипе. Кариотип – полный набор хромосом, характеризующийся у каждого вида их строго определенным числом и строением. Геномный уровень отличается высокой стабильностью. Он обеспечивает сложную систему взаимодействия генов. Результатом взаимодействия генов друг с другом и с факторами внешней среды является фенотип. 30. Доказательство генетической роли молекулы ДНК. Опыты Ф Гриффита. Эксперимент О Эвери, К Маклеода и М. Маккарти. Эксперимент А.Херши и М. Чейз. http://www.bio.bsu.by/genetics/files/2.molbiol_gene/molbiol_of_the_gene_03.pdf 31. Генный уровень организации наследственного материала. Эволюция понятия «ген». Строение молекулы ДНК. Репликация ДНК. https://ru.wikipedia.org/wiki/%D0%93%D0%B5%D0%BD 32. Классификация генов: структурные гены, гены-регуляторы, гены-модификаторы. Свойства генов (дискретность, стабильность, лабильность, полиаллелизм, специфичность, плейотропия). Свойства гена: Дискретность — несмешиваемость генов; Стабильность — способность сохранять структуру; Лабильность — способность многократно мутировать; Множественный аллелизм — многие гены существуют в популяции во множестве молекулярных форм; Аллельность — в генотипе диплоидных организмов только две формы гена; Специфичность — каждый ген кодирует свой признак; Плейотропия — множественный эффект гена; Экспрессивность — степень выраженности гена в признаке; Пенетрантность — частота проявления гена в фенотипе; Амплификация — увеличение количества копий гена. 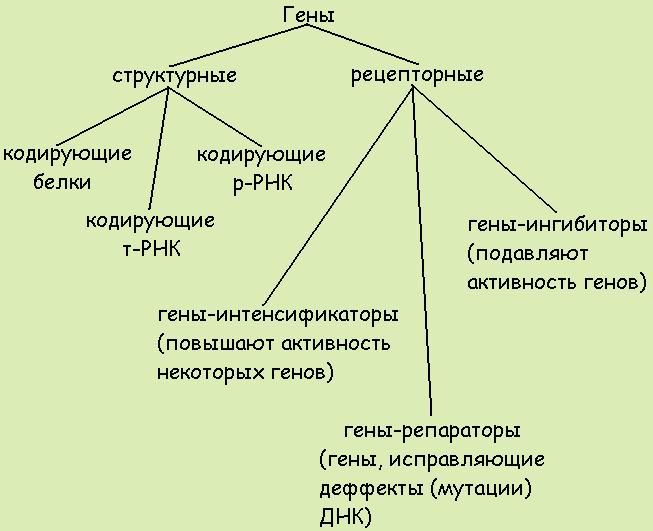 33. Экспрессия генов в процессе биосинтеза белка. Строение РНК. Виды РНК. Генетический код. Реализация наслед-ной инф-ии, заключенной в генотипе организма, - это сложный процесс, требующий тонкой регуляции для того, чтобы в кл разной тканевой принадлежности в опред-ное время в процессе развития орг-ма обеспечить синтез специф-их белков в необход. кол-ве. Основная масса генов, активно функционирующих в большинстве клеток организма на протяжении онтогенеза, - это гены, обеспечивающие синтез белков общего значения, тРНК и рРНК. Транскрибирование этих генов обеспечивается соединением РНК-полимеразы с их промоторами. Такие гены называются конститутивными. Другая группа генов, детерминирующих синтез специфических продуктов, в своем функционировании зависит от различных регулирующих факторов - регулируемые гены. Их активное функционирование, скорость и продолжительность транскрипции регулируются путем стимуляции или запрещения соединения РНК-полимеразы с промоторной областью гена. Регуляция экспрессии генов у прокариот: гипотеза оперона была предложена в 1961г. на примере лактозного оперона. Группа генов, расположенных в ДНК, управляется одним геном-опероном, образую оперон. В цитоплазме проходит стадия 7-9, на рибосомах 7-8. оперон работает по принципу обратной связи. 1-промотор, 2-инициатор, 3-оператор, 4-структурные гены, 5-терминатор, 6-ген-регулятор, 7-информационный репрессор, выключающий оператор, 8-фермент, 9-индуктор. В состав оперона входят 1,2- место первичного прикрепления РНК-полимераза, 3-вкл и выкл структурные гены, для считывания информации. Оперон – участок ДНК, на котором считывается иРНК, определяет синтез белка-фермента, ген-регулятор – содержит информацию для синтеза. 7 – блокирует 3, вступая с ним в химическое соединение информация не считывается, оперон не работает. Чтобы оперон заработал 7 должен быть связан с 9. Когда последние молекулы индуктора будут разрушены, освобождается репрессор, блокируется оператор (3). +Регуляция экспрессии генов у эукариот: гипотеза транскриптона – 1972г. Г.Георгиев. В клетке прокариота наследственный материал биосинтеза белка пространственно не разобщены, транскрипция и трансляция происходят почти одновременно. У эукариот 2 этапа, часть процесса проходит в ядре, часть – в цитоплазме и во времени их разделяют процессы созревания иРНК, из которой должны быть удалены некодирующие последовательности. Транскриптон – ед-ца транскрипции, состоящей из неинформационной и инф-ой зоны. 7-9 – в цитоплазме, 1-6 – в ядре. РНК — рибонуклеиновая кислота, имеет много общего со структурой ДНК, но отличается от нее рядом признаков: 1. углеводом РНК, к которому присоединяются пуриновые или пиримидиновые основания и фосфатные группы, является рибоза; 2. в состав РНК, как и в состав ДНК, входят азотистые основания аденин, гуанин и цитозин. Но РНК не содержит тимина, его место в молекуле РНК занимает урацил; 3. РНК — одноцепочечная молекула; 4. так как молекула РНК одноцепочечная, то правило Чаргаффа, установленное для ДНК, может не выполняться по равенству содержания оснований. РНК, присутствующие в клетках как протак и эукариот, бывают трех основных видов: матричные РНК (мРНК), рибосомные РНК (рРНК) и транспортные РНК (тРНК). Матричные РНК выполняют функцию матриц белкового синтеза. В ядре клеток эукариот содержится РНК четвертого типа гетерогенная ядерная РНК(гяРНК), которая является точной копией (транскриптом) соответствующей ДНК. Процесс транскрипции осуществляется в ядре на ДНК, гяРНК после созревания будет служить матрицей для синтеза белка в цитоплазме. Молекулы тРНК узнают в цитоплазме соответствующий триплет (кодон в мРНК) и переносят нужную аминокислоту к растущей полипептидной цепи. Узнавание кодона в мРНК осуществляется с помощью трех последовательных оснований в тРНК, называемыхантикодонами. Аминокислотный остаток может присоединятся к 3 — концу молекулы тРНК. Считают, что для каждой аминокислоты имеется, по крайней мере, одна тРНК. Молекула тРНК содержит около 75 нуклеотидов, ковалентно связанных друг с другом в линейную цепочку. рРНК— несколько молекул РНК, составляющих основу рибосомы. Основной функцией рРНК является осуществление процесса трансляции - считывания информации с мРНК при помощи адапторных молекул тРНК и катализ образования пептидных связей между присоединёнными к тРНК аминокислотами. 4. Генетический код и его свойства +Генетический код, система зашифровки наследственной информации в молекулах нуклеиновых кислот, реализующаяся у животных, растений, бактерий и вирусов в виде последовательности нуклеотидов. В природных нуклеиновых кислотах — дезоксирибонуклеиновой (ДНК) и рибонуклеиновой (РНК) — встречаются 5 распространённых типов нуклеотидов (по 4 в каждой нуклеиновой кислоте), различающихся по входящему в их состав азотистому основанию. В ДНК встречаются основания: аденин (А), гуанин (Г), цитозин (Ц), тимин (Т); в РНК вместо тимина присутствует урацил (У). Кроме них, в составе нуклеиновых кислот обнаружено около 20 редко встречающихся (т. н. неканонических, или минорных) оснований, а также необычных сахаров. Т. к. количество кодирующих знаков Г. к. (4) и число разновидностей аминокислот в белке (20) не совпадают, кодовое число (т. е. количество нуклеотидов, кодирующих 1 аминокислоту) не может быть равно 1. 34. Регуляция экспрессии генов прокариот и эукариот. Экспрессия генов — это реализация заложенной в них информации, то есть синтез РНК и белков. Другими словами, под экспрессией генов понимают их активность. В клетках живых организмов экспрессия генов регулируется: одни гены могут быть реализованы, другие — нет. Причем регуляция может осуществляться на разных этапах: может выполняться или нет транскрипция, из пре-мРНК в результате альтернативного сплайсинга могут образовываться разные мРНК, может блокироваться трансляция и др. У эукариот, обладающих отграниченным от цитоплазмы ядерным содержимым и более сложным геномом, регуляция экспрессии генов намного разнообразнее и сложнее, чем у прокариот. Регуляция экспрессии генов у прокариот У прокариот пока молекула РНК синтезируется на участке ДНК, она тут же может транслироваться (начиная с уже синтезированного конца). Поэтому у них регуляция экспрессии (активности) генов осуществляется почти исключительно на уровне ДНК, так как в РНК часто невозможно внести какие-нибудь изменения до ее трансляции. В 1961 г. Жакобом и Моно была предложена модель оперона как системы регуляции генов у бактерий. Оперон состоит из промотора, оператора, структурных генов оперона (их может быть разное количество) и терминатора. В области промотора прикрепляется фермент РНК-полимераза. В области оператора присоединяется белок-репрессор, который кодируется отдельно отстоящим от оперона геном-регулятором (может быть сцеплен со своим опероном, а может находиться на расстоянии). Если белок-репрессор соединяется с оператором, то транскрипция всех структурных генов оперона становится невозможной, так как РНК-полимераза не может перемещаться по цепи ДНК. В свою очередь активность белка-репрессора может блокироваться определенным для него низкомолекулярным соединением — индуктором (тем или иным питательным веществом бактерий). В результате взаимодействия с индуктором белок-репрессор видоизменяется и уже не может присоединиться к оператору своего оперона. В этом случае гены оперона экспрессируются (т. е. на них идет синтез). Бывает обратная ситуация, когда индуктор активирует белок-репрессор. Таким образом, в зависимости от того, какие индукторы находятся в цитоплазме, у прокариот экспрессируются те или иные генные группы. Вышеописанный механизм экспрессии генов относится к негативной регуляции, так как гены транскрибируются, если они не выключены репрессором. И наоборот: не транскрибируются, если выключены. Кроме негативной регуляции у бактерий существует также позитивная. В этом случае вместо белка-репрессора действие оказывает белок-активатор. На эти белки также действуют индукторы, активируя или инактивируя их. Также у прокариот были выявлены опероны, которые актируются двумя регуляторными белками, соединенными друг с другом. Регуляция экспрессии генов у эукариот У многоклеточных организмов в клетках разных тканей экспрессируются разные гены, т. е. для эукариот характерна дифференциальная экспрессия. У эукариот, также как и у прокариот, существуют регуляторные белки с похожим механизмом действия. При этом для эукариот не характерна регуляция по типу оперона. Цистроны (транскрибируемые участки) эукариот обычно содержат по одному гену. (Это не касается геномов хлоропластов и митохондрий.) Кроме регуляторных белков, взаимодействующих с ДНК, у эукариот существуют и другие способы регуляции экспрессии генов. Конденсация и деконденсация хроматина. Это наиболее универсальный метод регуляции транскрипции. Когда нужно экспрессировать определенные гены, хроматин в этом месте деконденсируется. Альтернативные промоторы. У гена может быть несколько промоторов, каждый из которых начинает транскрипцию с разных его экзонов в зависимости от типа клетки. В конечном итоге будут синтезированы разные белки. Метилирование и деметилирование ДНК. Метилирование ДНК происходит в регуляторных областях гена. Метилируется цитозин в последовательности ЦГ, после чего ген инактивируется. При деметилировании активность гена восстанавливается. Процесс регулируется ферментом метилтрансферазой. Гормональная регуляция. При гормональной регуляции гены активируются в ответ на внешний химический сигнал (поступление в клетку определенного гормона). Этот гормон запускает те гены, которые имеют специфические последовательности нуклеотидов в регуляторных областях. Геномный импринтинг. Это малоизученный способ регуляции экспрессии генов у эукариот. Он возможен только у диплоидных организмов и выражается в том, что активность генов зависит, от какого из родителей они были получены. Выключение генов осуществляется путем метилирования ДНК. Альтернативный сплайсинг. Это регуляция на уровне процессинга. При альтернативном сплайсинге порядок сшивки экзонов может быть различным. Отсюда следует, что на основе одной и той же нуклеотидной последовательности ДНК могут быть синтезированы разные белки. Хотя их отличие друг от друга будет в основном заключаться лишь в разных сочетаниях одних и тех же аминокислот. Тканеспецифическое редактирование РНК также протекает на уровне процессинга. Выражается в замене отдельных нуклеотидов в РНК в определенных тканях организма. Кроме того, у эукариот иРНК часто не подвергается процессингу вообще (а распадается) или подвергается с задержкой. Это токже можно рассматривать как способ регуляции экспрессии генов. Регуляция стабильности иРНК. У эукариот существует регуляция и на уровне трансляции, когда готовые иРНК не «допускаются» к рибосомам или разрушаются. Другие же иРНК могут дополнительно стабилизироваться для многократного использования. Посттрансляционная модификация белка. Чтобы молекула полипептида превратилась в активную молекулу белка, в ней должны произойти различные модификации определенных аминокислот, должны быть сформированы вторичная, третичная и возможно четверичная структуры. На этом этапе также можно повлиять на реализацию генетической информации, например, не дав молекуле сформироваться. Риборегуляторы. Были обнаружены РНК, выполняющие регуляторные функции путем ослабления работы отдельных генов. Для высокоорганизованных животных отмечается существование надклеточного уровня регуляции экспрессии генов. |